Как ДНК становится трехмерной: структура хроматина
Александра Галицына
Приглашенная лекция для ШМТБ 2020





Классические представления о ДНК
- ДНК - это носитель генетической информации, линейно упорядоченная молекула биополимера нуклеиновых кислот.

Первое доказательство линейности ДНК
- Ранние эксперименты Бензера
на E. coli и T4 бактериофаге:
Benzer, 1950-1960
https://slideplayer.com/slide/1400263/

Плашка с бактериями,
появление бляшек при заражении фагом
Первое доказательство линейности ДНК
- Ко-трансфекция двумя мутантными вирусами:
Benzer, 1950-1960
https://slideplayer.com/slide/1400263/
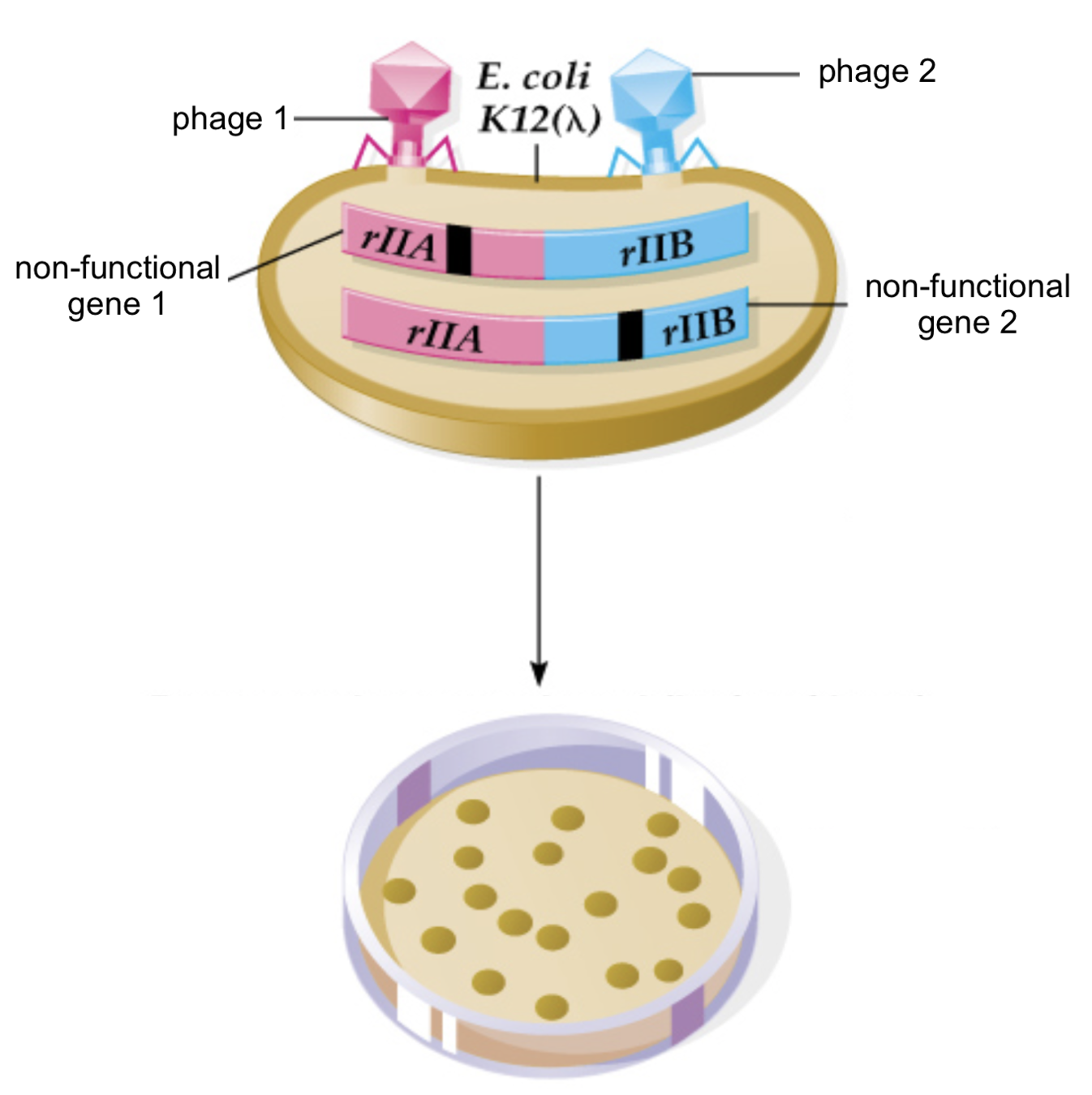
Появление бляшек (заражение есть)
Появление бляшек (заражения нет)
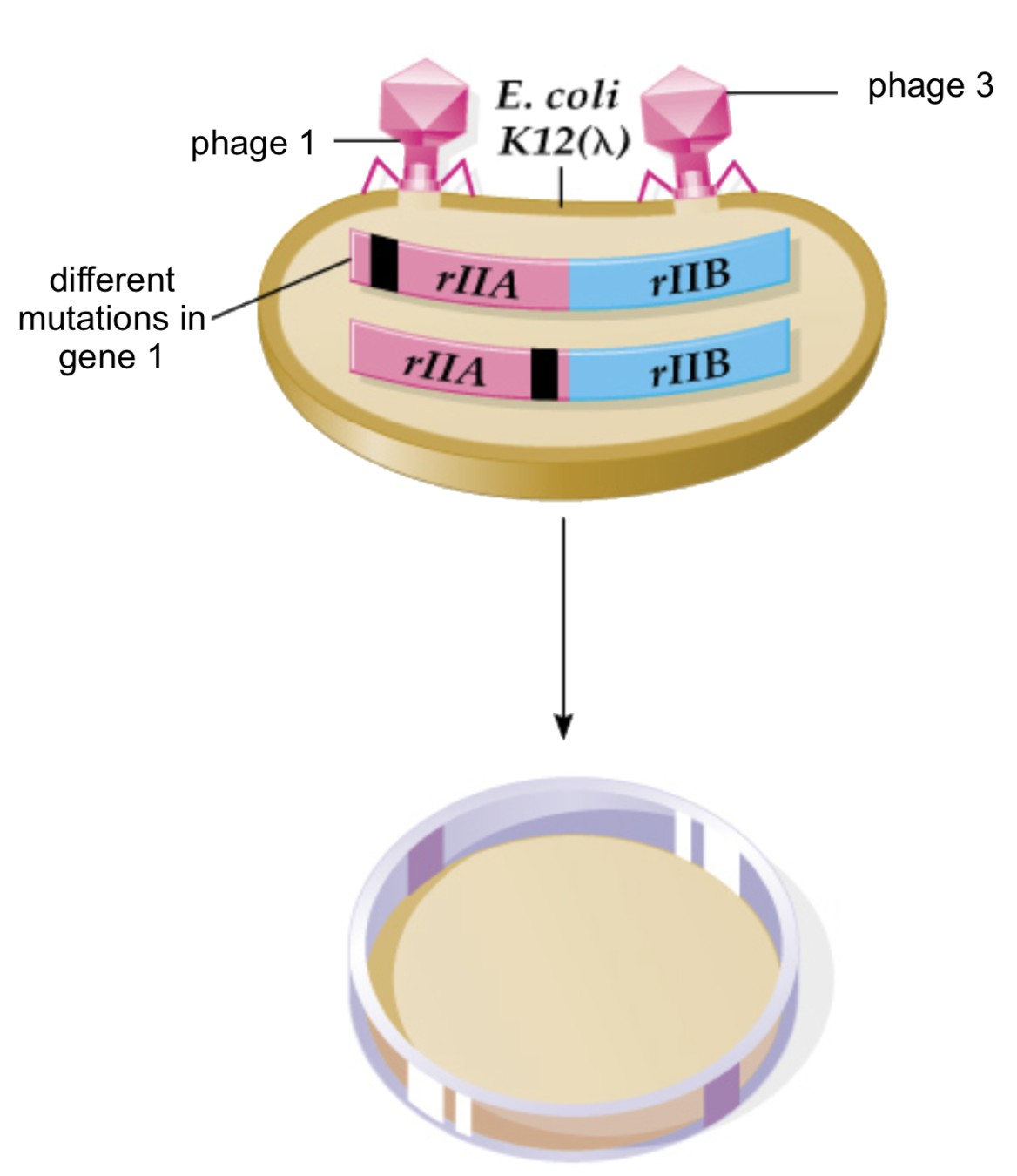
Первое доказательство линейности ДНК
- Генетическая карта бактериофага:
Benzer 1961
О - нет заражения, |- заражение есть
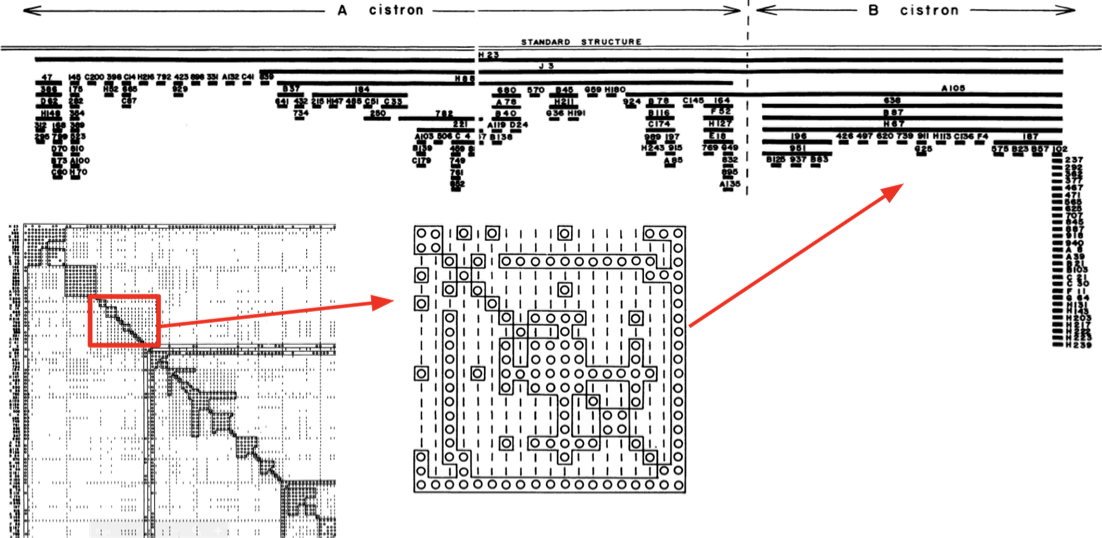
Упорядоченные мутации фага
ДНК как двойная спираль
- Уотсон, Крик и Франклин: рентгеноструктурный анализ (1953)

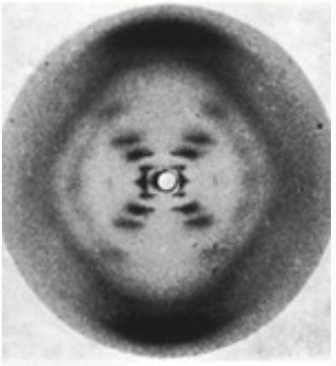
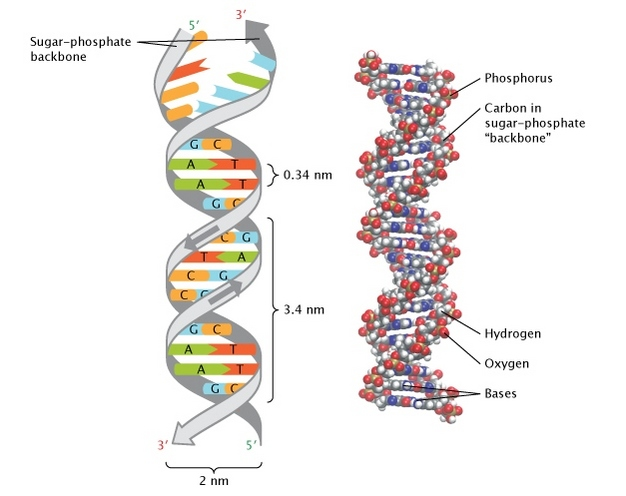
Pray Nature Education 2008
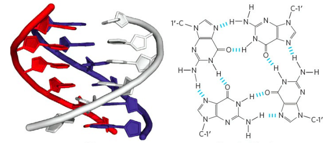


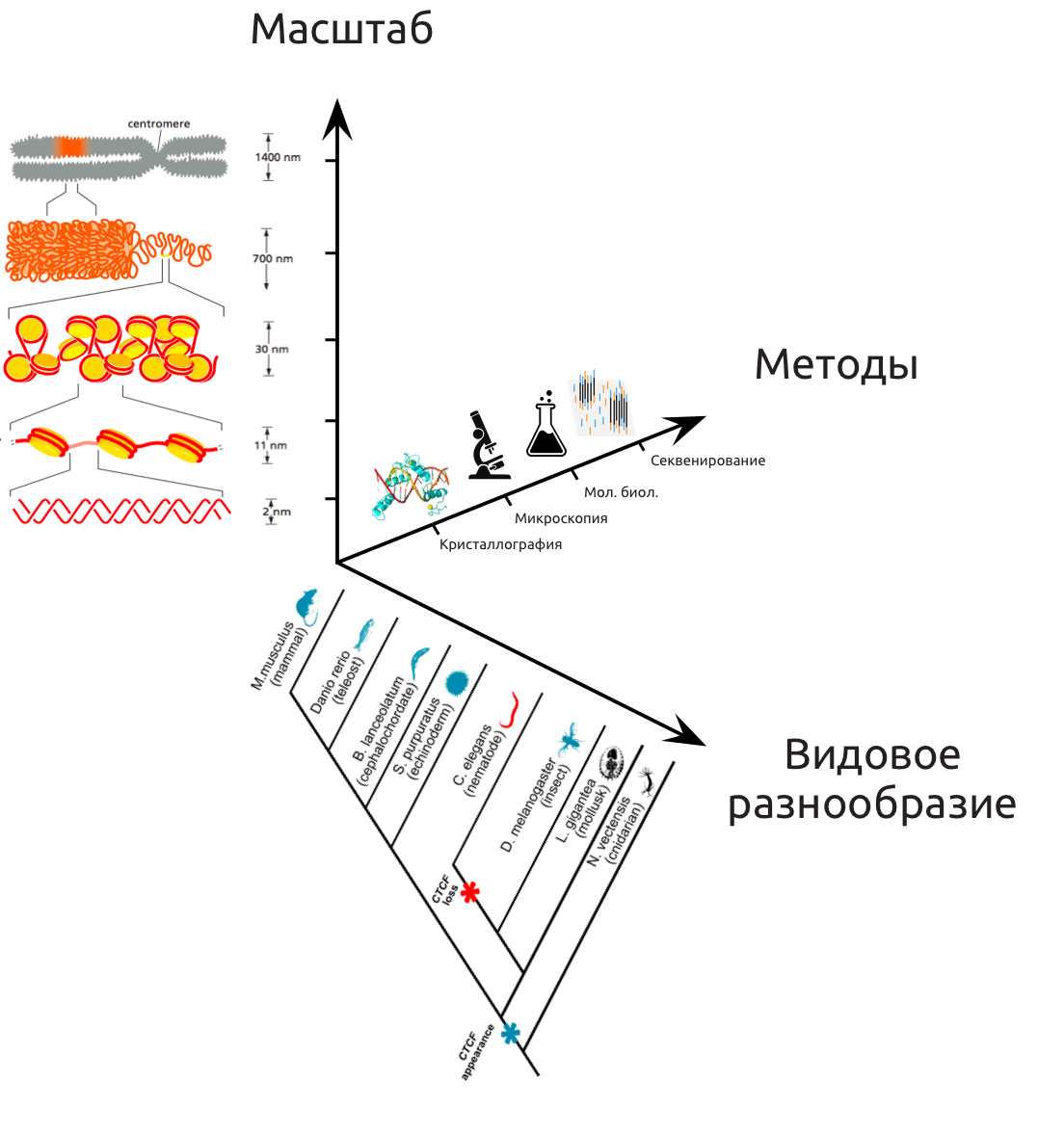
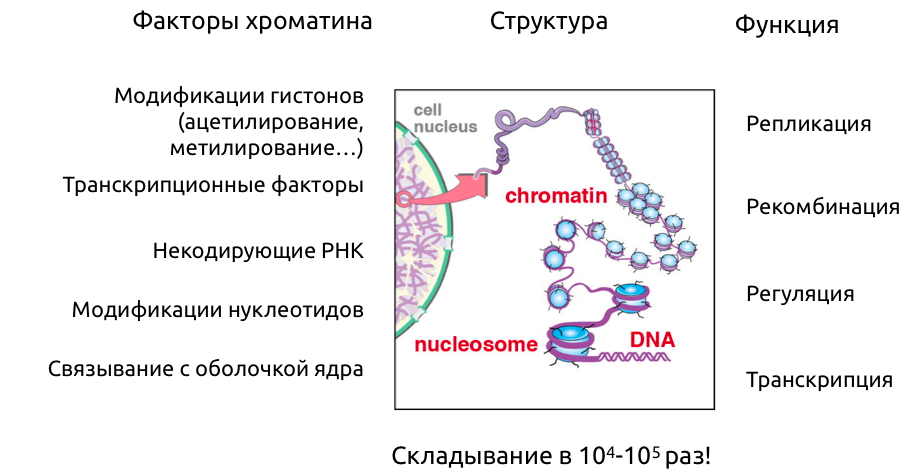
Уровни организации хроматина
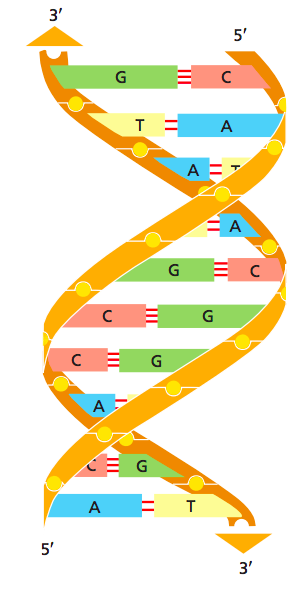
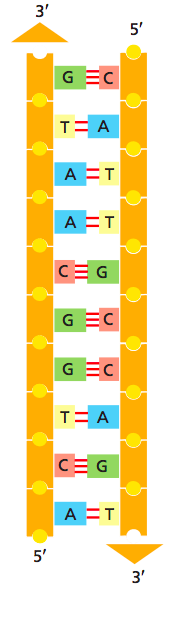
Первичная структура
Вторичная структура
A-форма
спирали
B-форма
Z-форма
G-квадруплекс
ДНК-шпилька
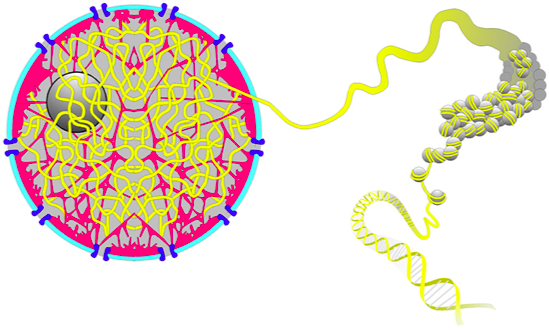
Хроматин - комплекс ДНК и белков
ДНК образует более сложные структуры за счет связывания белков. В первую очередь, это гистоновые белки (у эукариот):
Alberts 2015 "Molecular Biology of the Cell" 6th edition
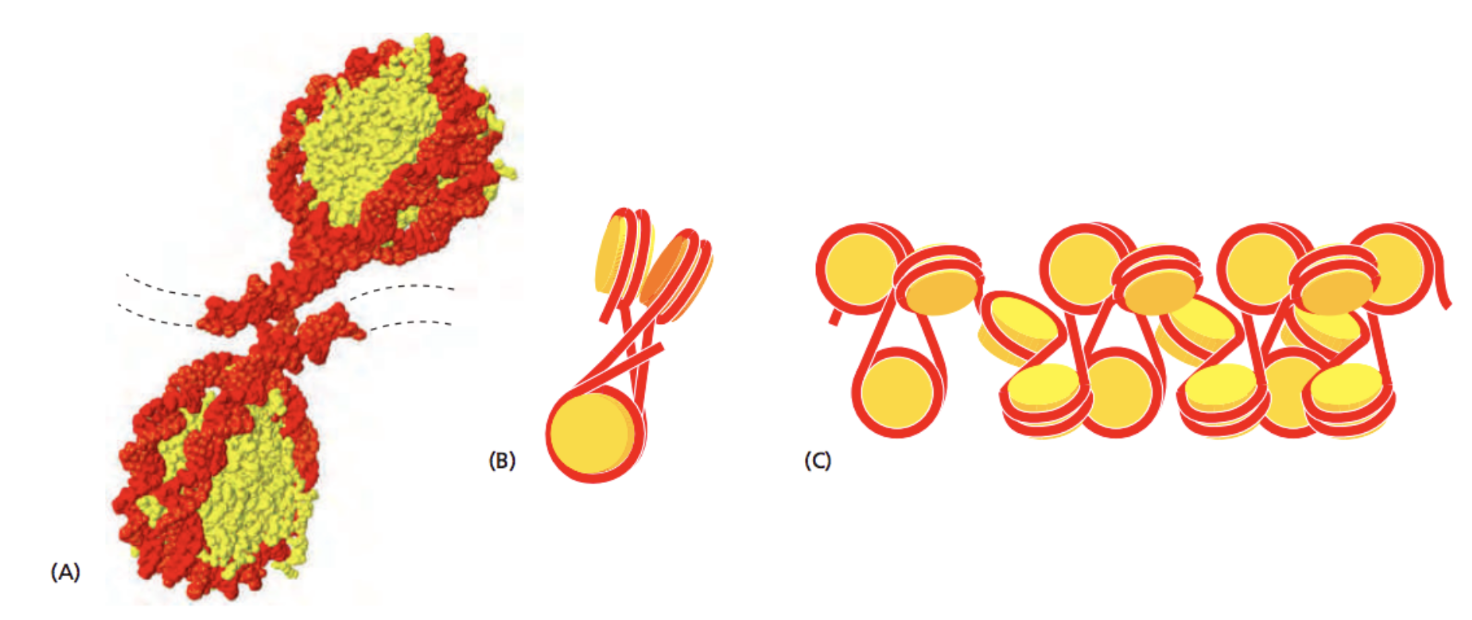
Хроматин - комплекс ДНК и белков
Связывание белковых факторов транскрипции:
Robinson, 2016
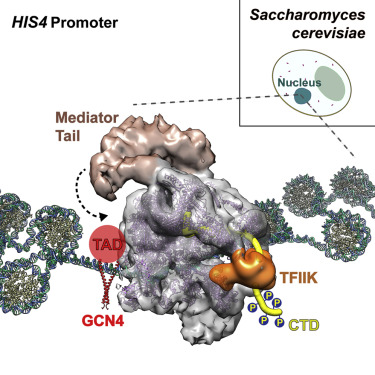
Регуляторы экспрессии генов: энхансеры
Для активации экспрессии необходима сборка белкового комплекса на специальных регуляторных последовательностях ДНК - энхансерах.
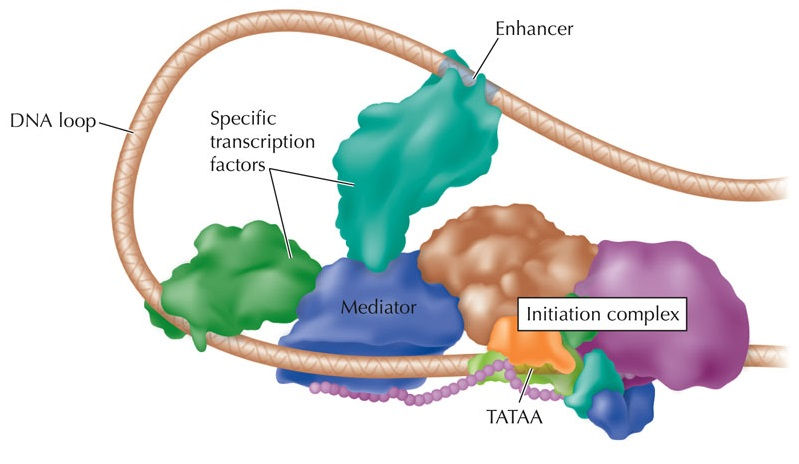
Энхансер
Промотор
Как привсти во взаимодействие промотор и энхансер?
Энхансер может располагаться на десятках тысяч нуклеотидов от промотора подрегулируемого гена.
Гипотетические механизмы:
Dean 2006
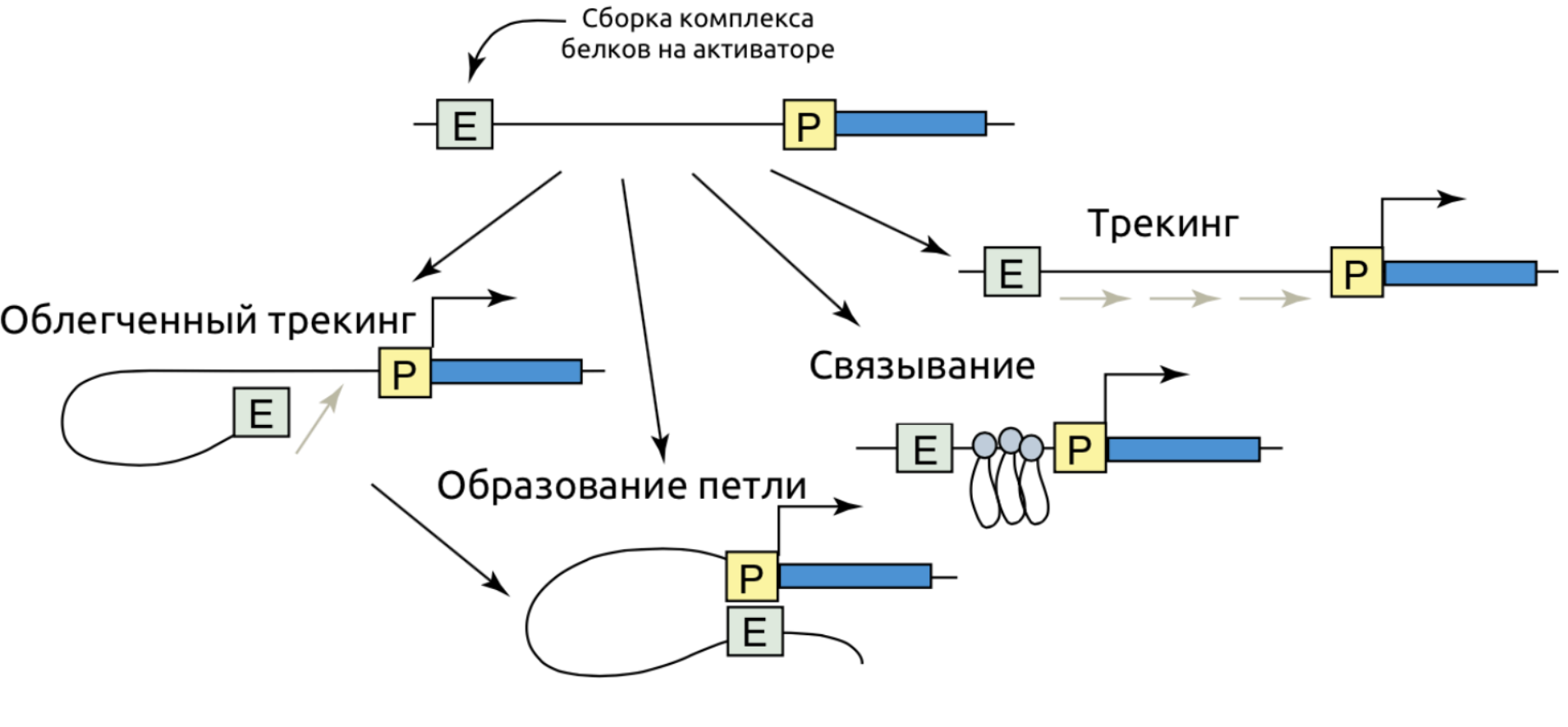
Регуляторные сети энхансеров
Один ген может регулироваться несколькими энхансерами, один энхансер может регулировать несколько генов:
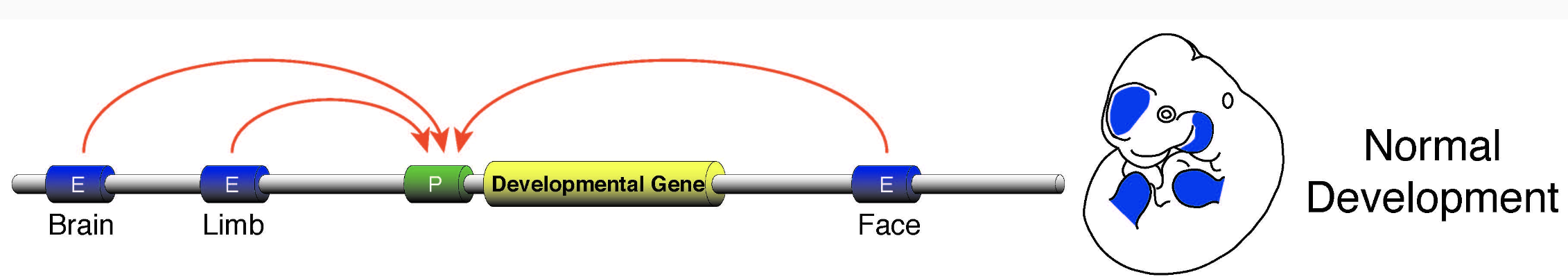
Регуляторные сети энхансера
Разнообразие "дальней" регуляции позволяет генерировать всевозможные фенотипы клеток многоклеточного организма:
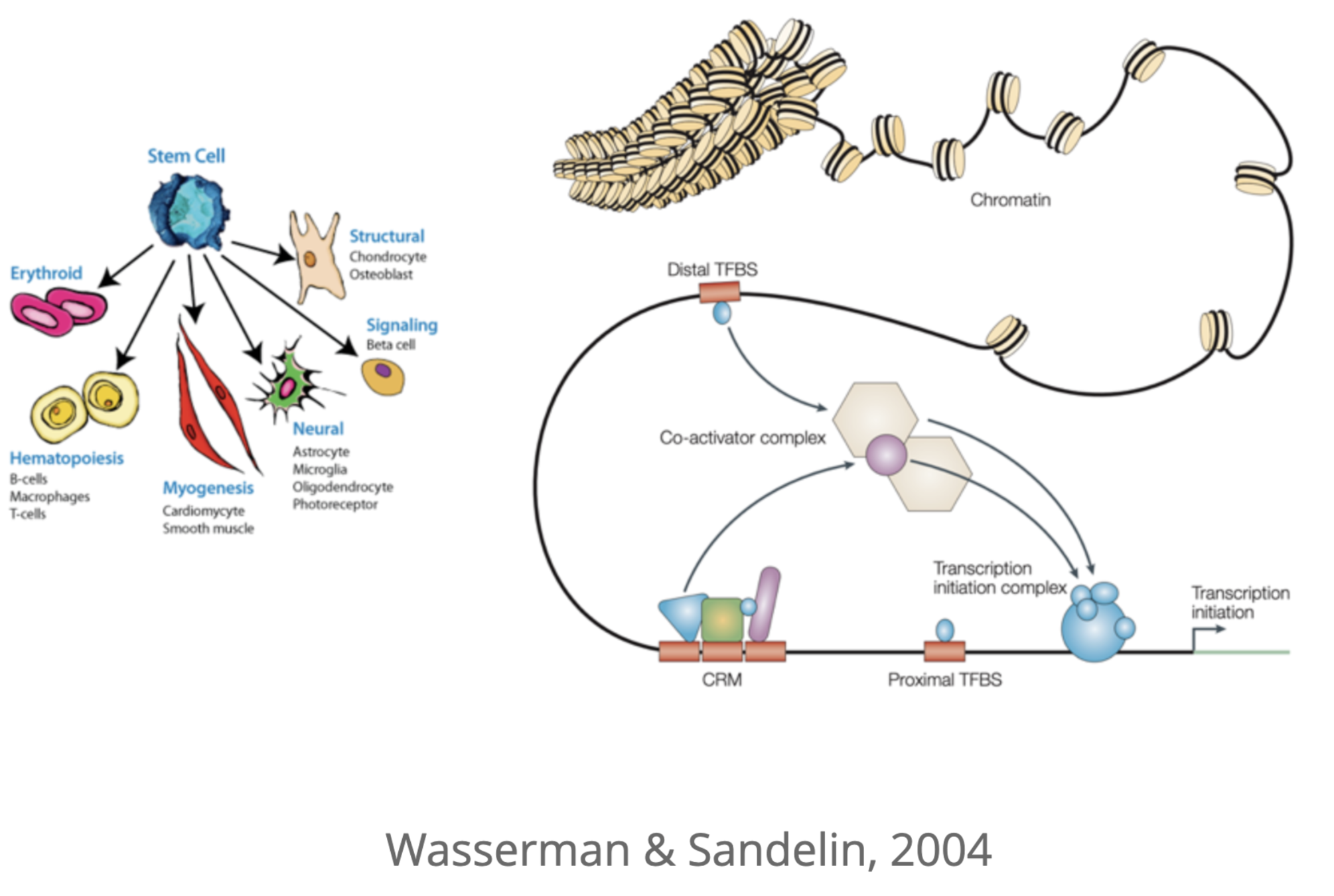
Подходы к исследованию хроматина и "дальней" регуляции
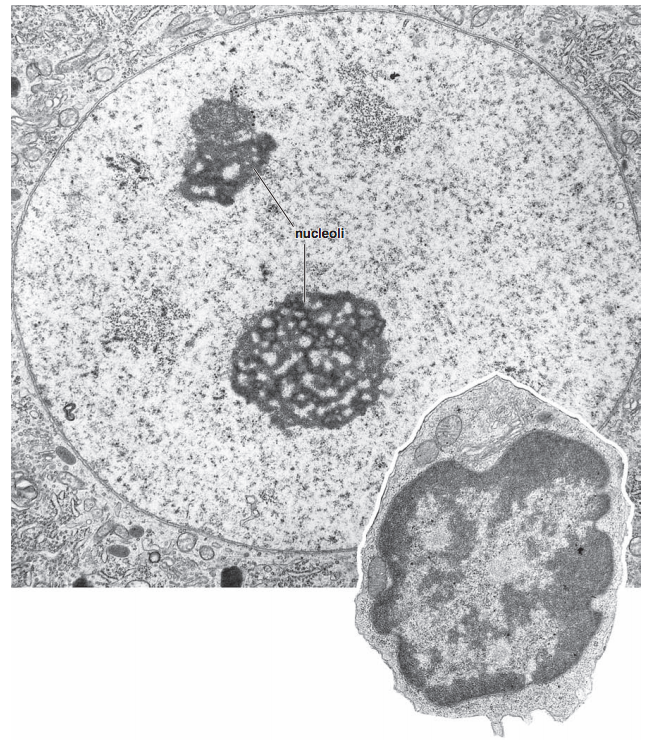
Микроскопия ядра
Ros 2006 "Histology Atlas with Correlated Cell and Molecular Biology"
Два подхода: микроскопия и молекулярная биология




FISH-микроскопия
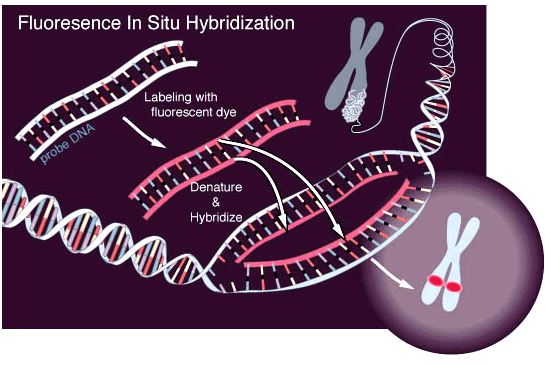
Fluorescent in situ hybridization
FISH-микроскопия
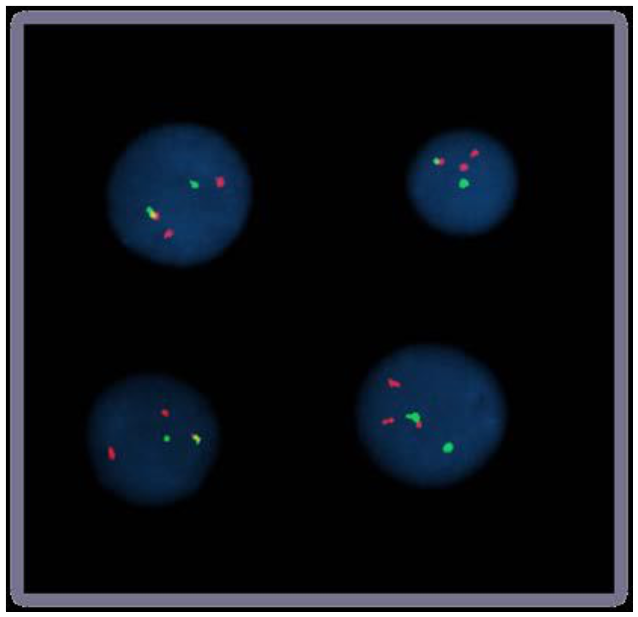
Bolzer et al., PLoS Biol. 2005
Speicher & Carter 2005 Nature
Fluorescent in situ hybridization
Фиксация конформации хромосом
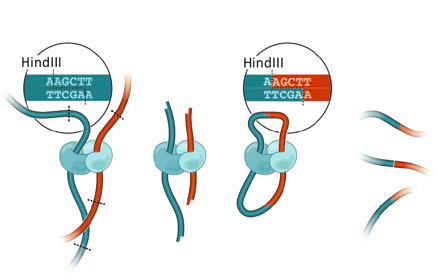
Фиксация формальдегидом
Рестрикция ДНК
Лигирование
Очистка ДНК
Библиотека ДНК-ДНК контактов
3C: Dekker et al. 2002 Science
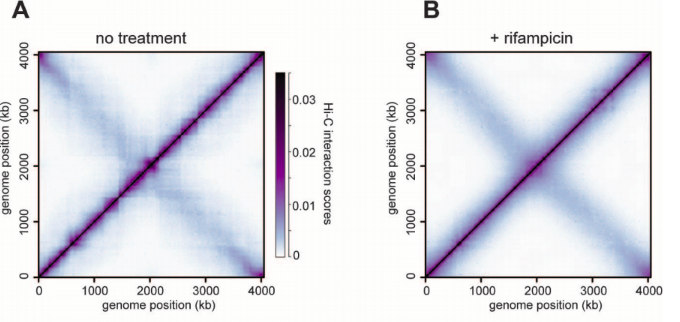
Hi-C: фиксация конформации хромосом + секвенирование
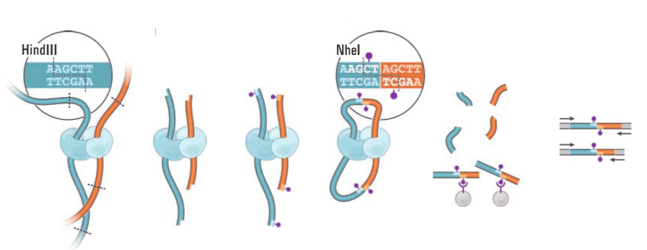
Фиксация формальдегидом
Рестрикция ДНК
Лигирование
Очистка ДНК
Секвенирование
Картирование
Lieberman-Aiden et al. 2009 Science
Лигирование
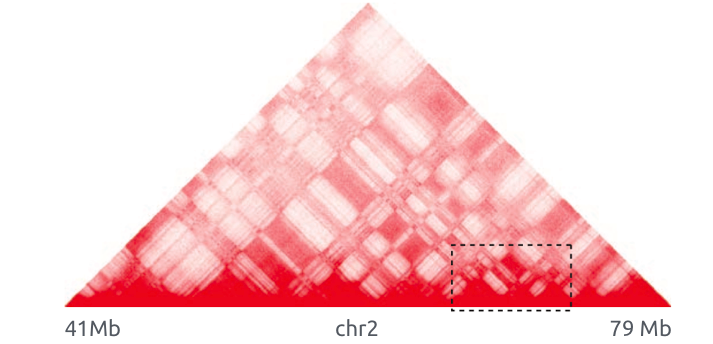
Теплокарта ДНК-ДНК контактов
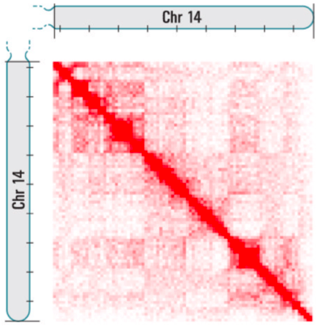
Lieberman-Aiden et al. 2009 Science
Цвет: частота взаимодействий регионов ДНК
Теплокарта ДНК-ДНК контактов
Lieberman-Aiden et al. 2009 Science
Цвет: частота взаимодействий регионов ДНК
Карта попарных взаимодействий
студентов ШМТБ-2020

Познакомились на летней школе, ботают вместе молбиол
Цвет: количество сообщений в Telegram
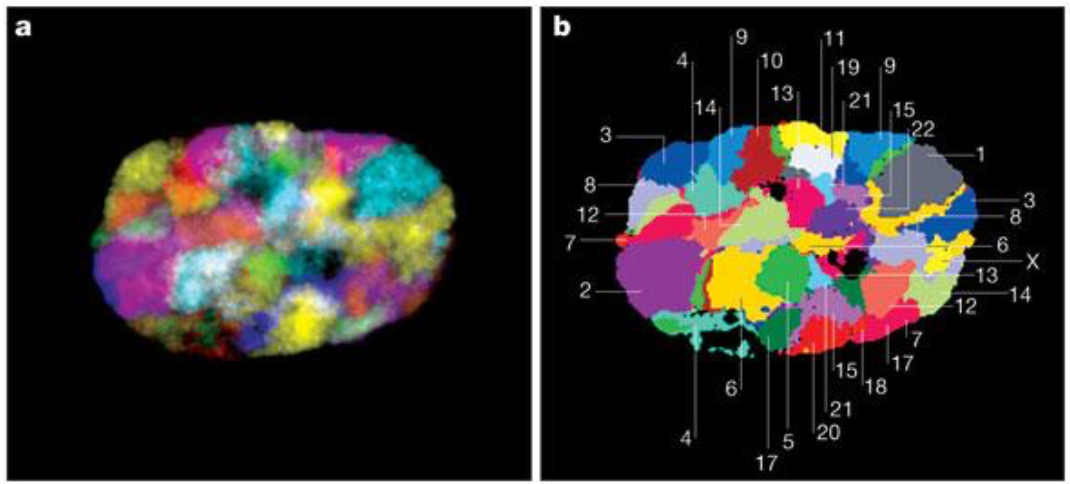
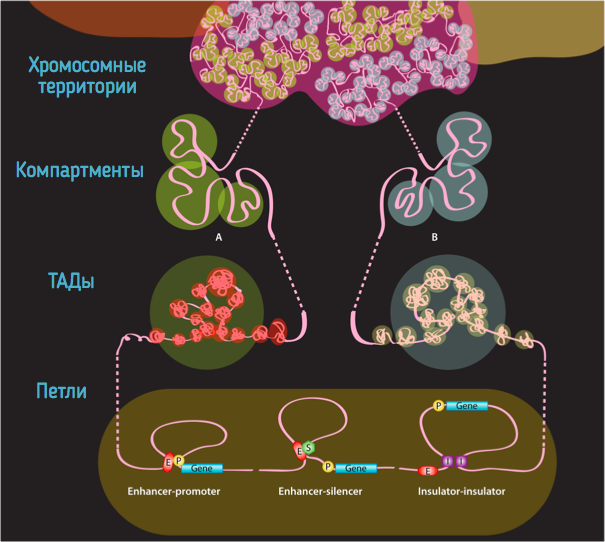
Хромосомные территории
Bonev et al. 2016 Nature Reviews
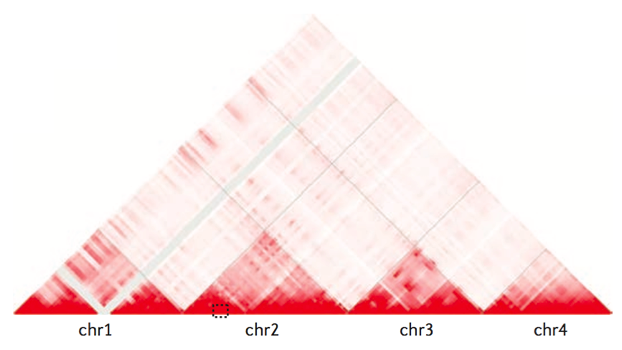
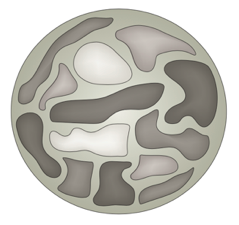
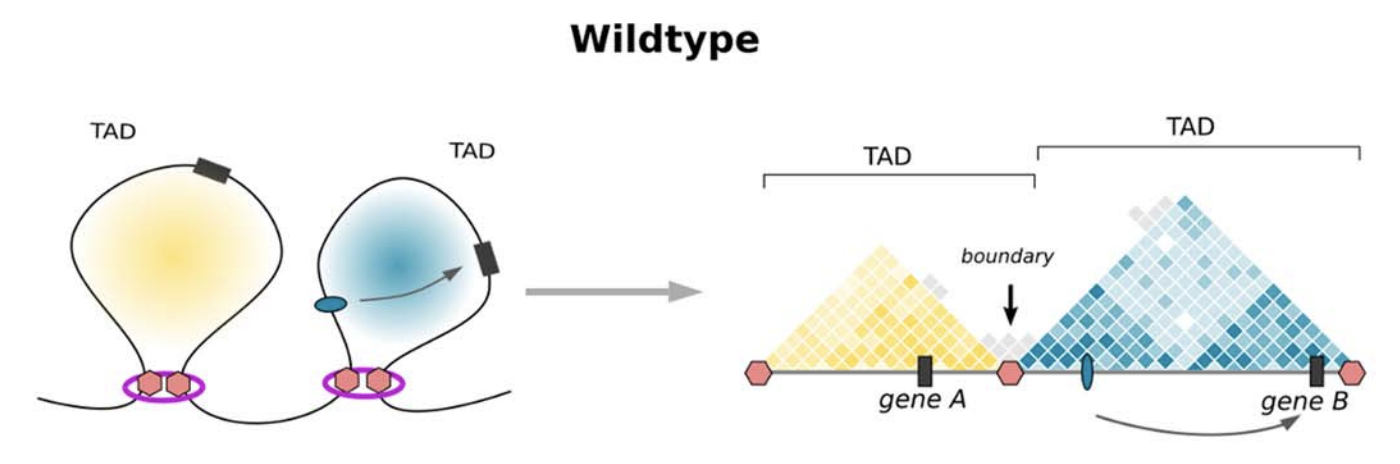
Топологически ассоциированные домены (ТАДы)

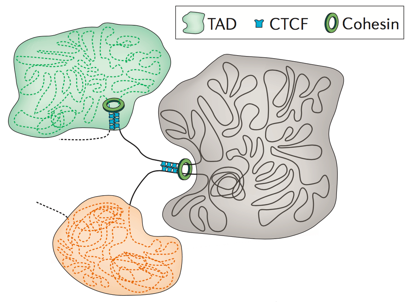
Bonev et al. 2016 Nature Reviews
Компартменты
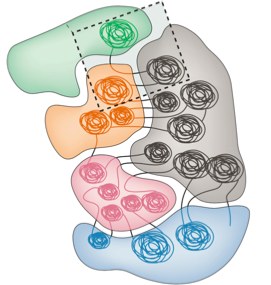
Bonev et al. 2016 Nature Reviews
Петли
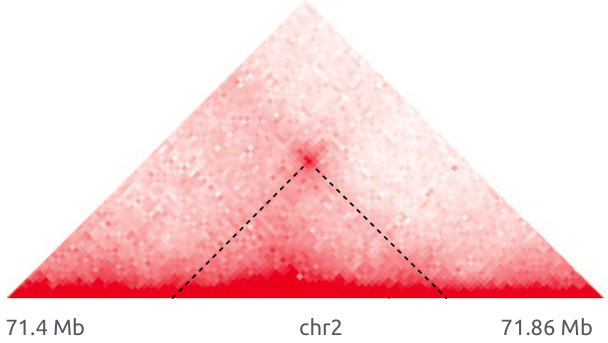
Архитектурные петли
Промотор-энхансерные петли
Polycomb-петли
Bonev et al. 2016 Nature Reviews
Уровни иерархии хроматина
Экструзия петель - основная гипотеза формирования структуры хроматина
MirnyLab Youtube channel
Упрощенная визуализация ключевых компонентов:
ДНК, экструдера (когезин) и барьерного элемента (CTCF)
Модель выпетливания
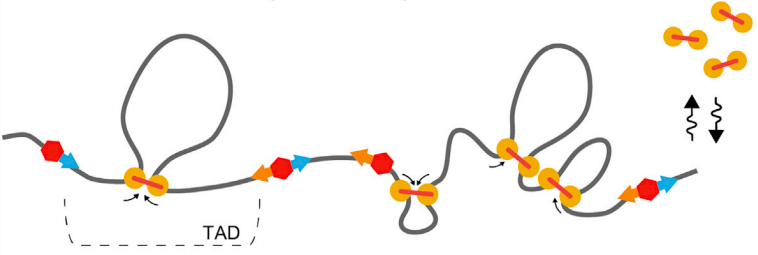
Fudenberg et al. 2016 Cell Reports

когезин - выпетливающий фактор
CTCF - граничный элемент
Архитектурный белок хроматина CTCF
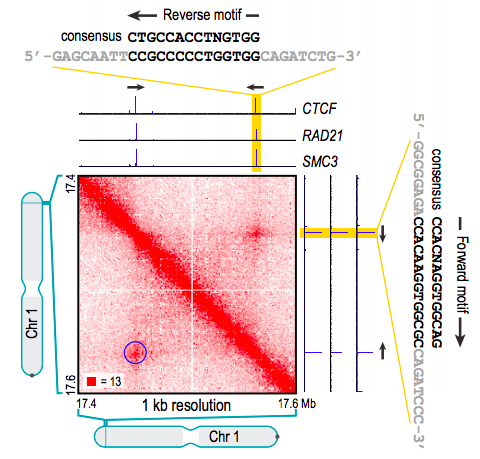
Rao et al. 2014 Cell
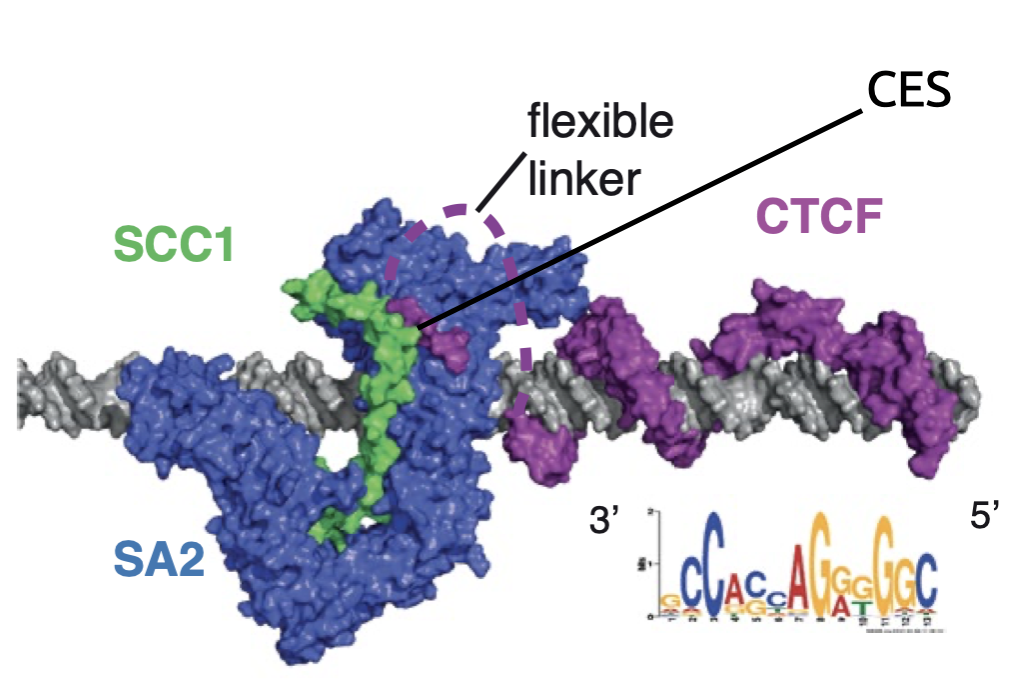
Ganji et al. 2018 Science
Ganji et al. 2018 Science
Прямое подтверждение экструзии
Модель активной компактизации хромосом
Mirny Lab Youtube channel
ДНК как активно живущий "город", наполненный активными машинками-экструдерами
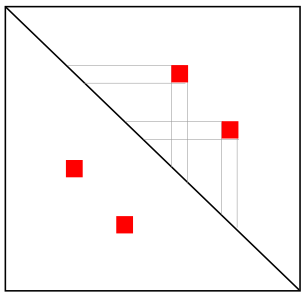
Квиз: реконструкция модели по карте Hi-C
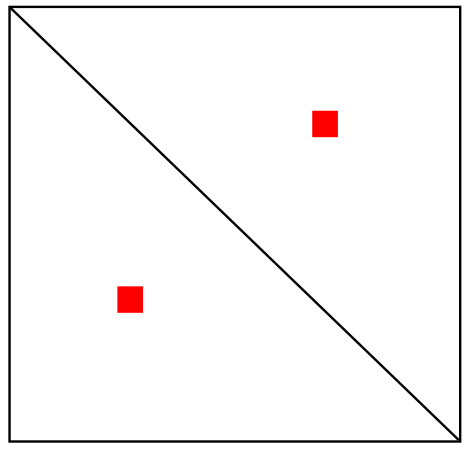

Петля
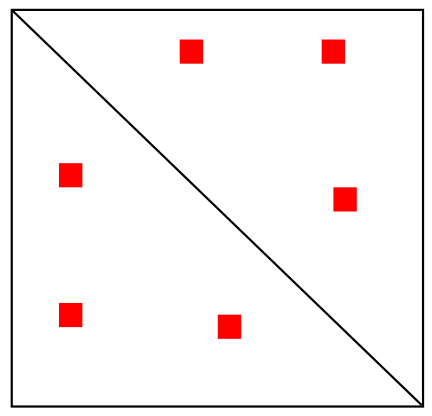

Реконструкция модели по карте Hi-C
Несколько петель
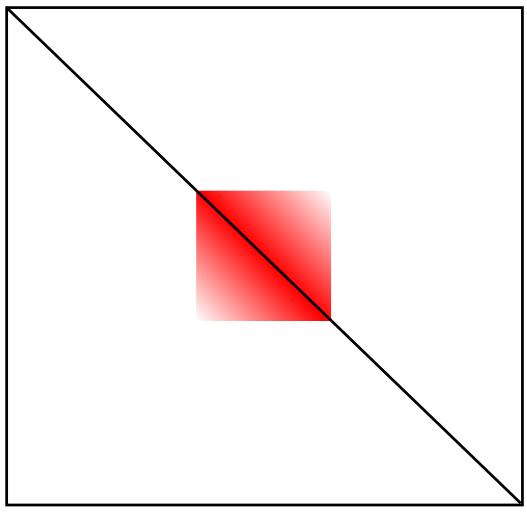

Реконструкция модели по карте Hi-C
Топологически-ассоциированный домен (ТАД)
Более экзотическая структура...
шпилька

Настоящие шпильки хроматина
Caulobacter crescentus
(бактерия)

Настоящие шпильки хроматина
Caulobacter crescentus
Псевдоузел
Редкий случай
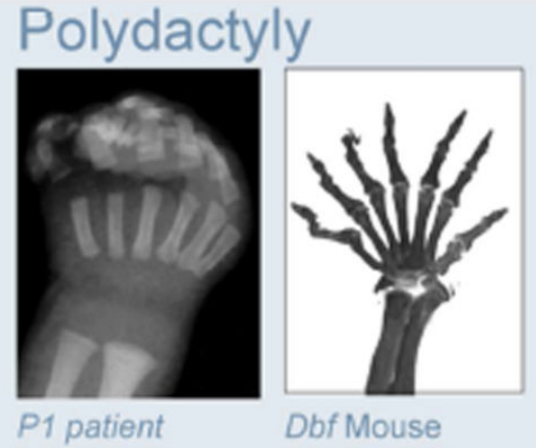
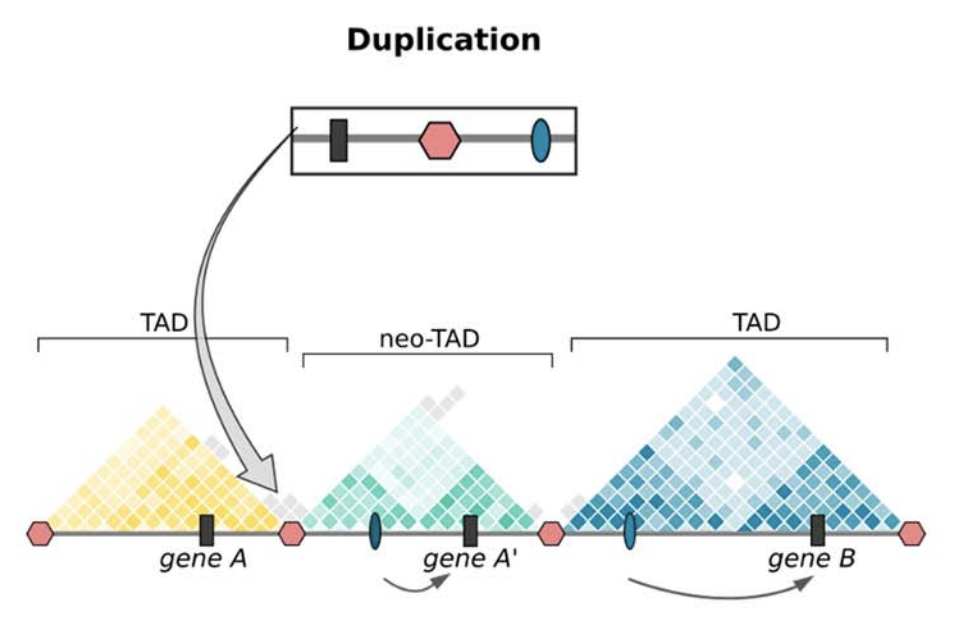
Клинические примеры
Anania and Lupiáñez, 2020; Lupiáñez et al. 2015
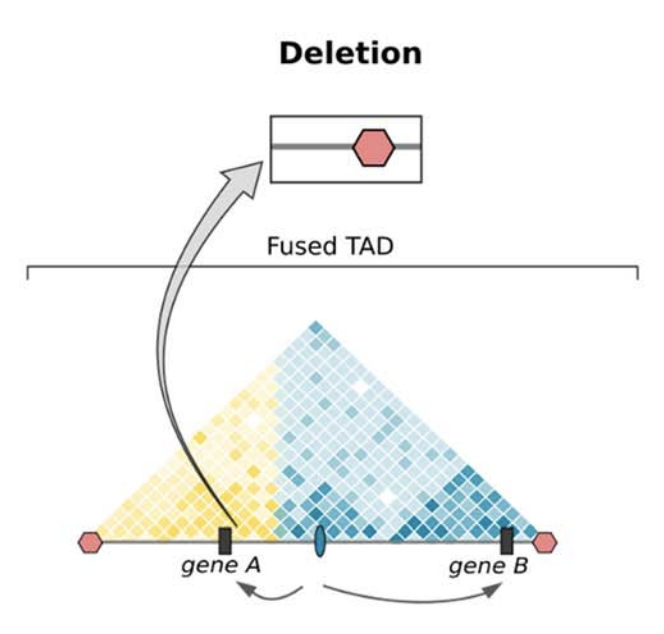
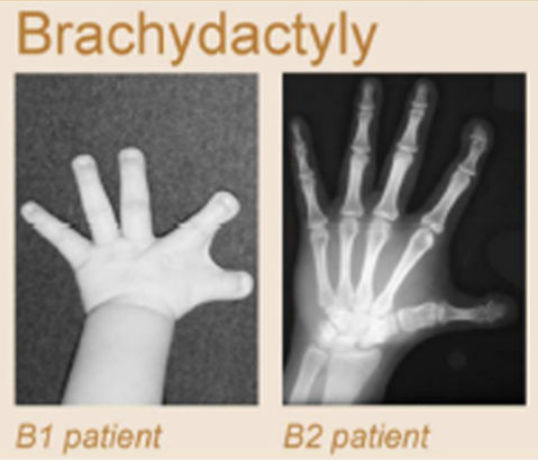
Клинические примеры
Anania and Lupiáñez, 2020; Lupiáñez et al. 2015
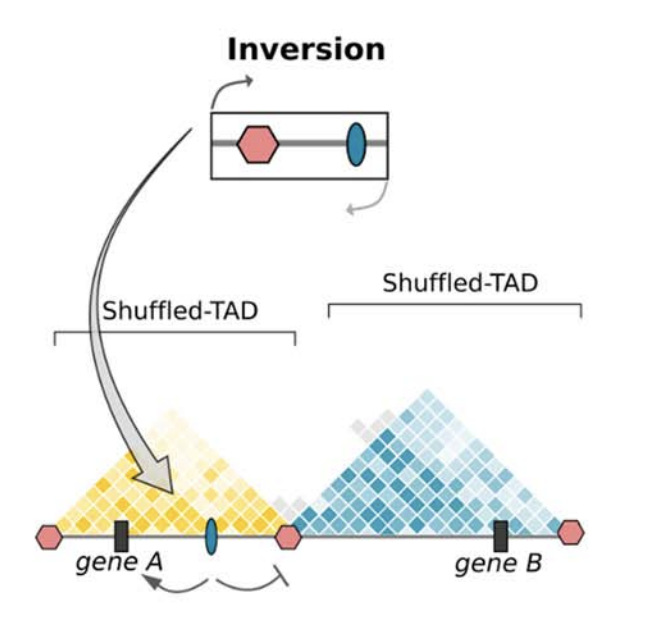
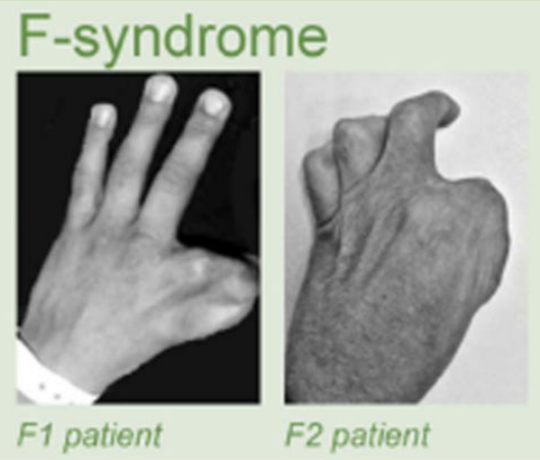
Клинические примеры
Anania and Lupiáñez, 2020; Lupiáñez et al. 2015
Take-home message
Человек: 2 м ДНК в 10 мкм ядро
100-этажный дом в рисовое зерно

Структура хроматина для ШМТБ 2020
By agalicina
Структура хроматина для ШМТБ 2020
Как ДНК становится трехмерной: структура хроматина Классическая генетика научила нас воспринимать геном как линейную ДНК с последовательно закодированными генами. Однако она не учитывает, что 2 метра ДНК упакованы в крошечное ядро, менее 10 микрометров в диаметре! Есть ли шанс, что это происходит случайно, без сложного молекулярного механизма, управляющего этим процессом? Недавние исследования показали, что ядро ведет себя скорее как оживленный город, где несколько типов молекулярных машин активно упаковывают ДНК и управляют экспрессией генов.




