









Jesse Bloom PRO
Scientist studying evolution of proteins and viruses.
(brainstorming slides for Bloom and Baker labs)
These slides: https://slides.com/jbloom/bloom-baker-dms-for-ab-design
The Bloom lab has measured how all mutations to viral entry proteins affect the function of these proteins and their neutralization by antibodies. These data:
These slides first provide some brief background on the topic and prior work by the Bloom lab. They then provide details on specific datasets that might help with the above goals.
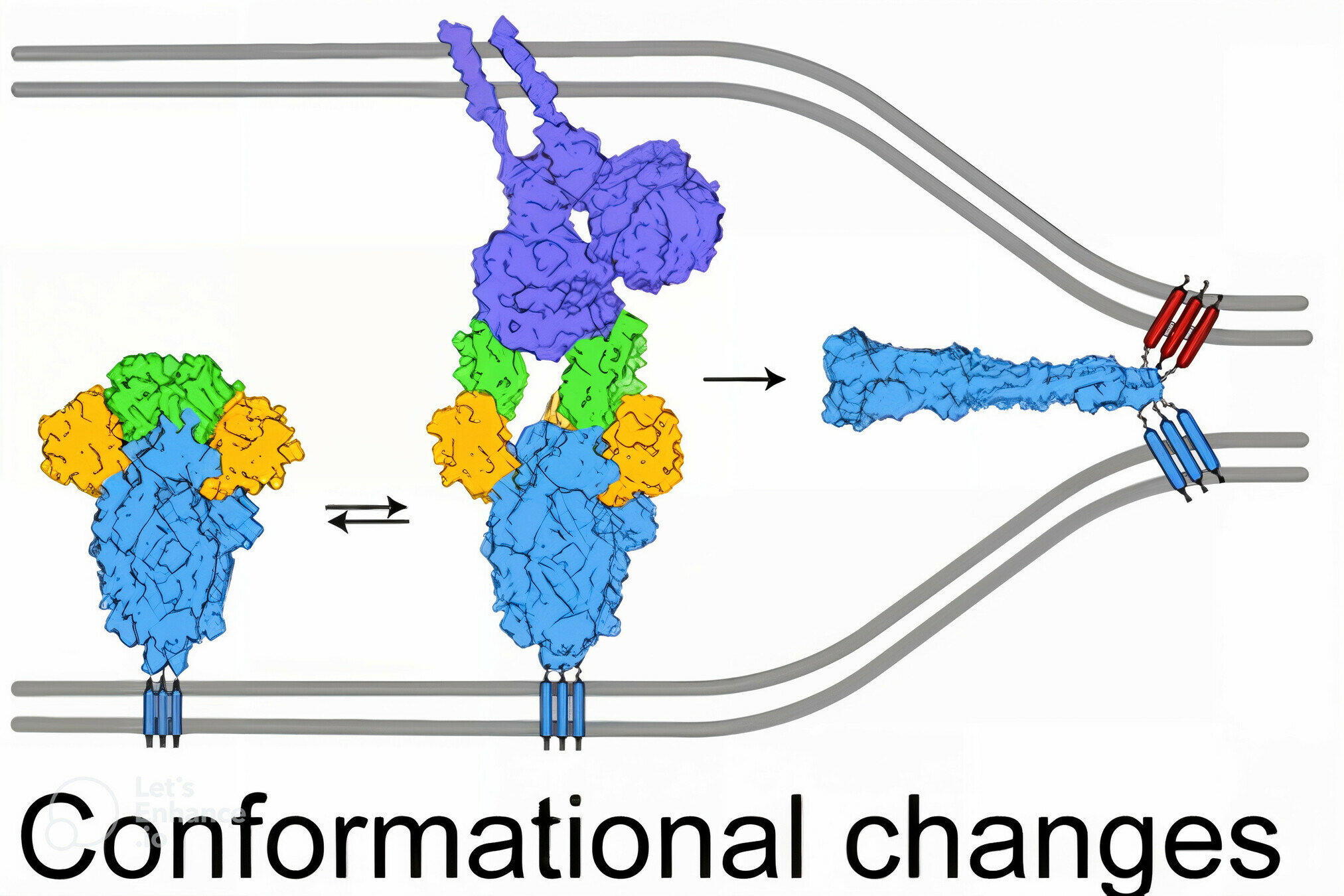
viral membrane
cell membrane
SARS-CoV-2 spike
spike conformational change
Image adapted from here
ACE2
antibody
Image adapted from here
In USA, RSV is the leading cause of infant hospitalization. Infants hospitalized with RSV receive supportive care (eg, oxygen, ventilation, fluids) and usually recover (~0.1% in-hospital case-fatality rate)
In developing world where supportive care not available, RSV is a leading cause of infant mortality (~100,000 infant deaths per year)
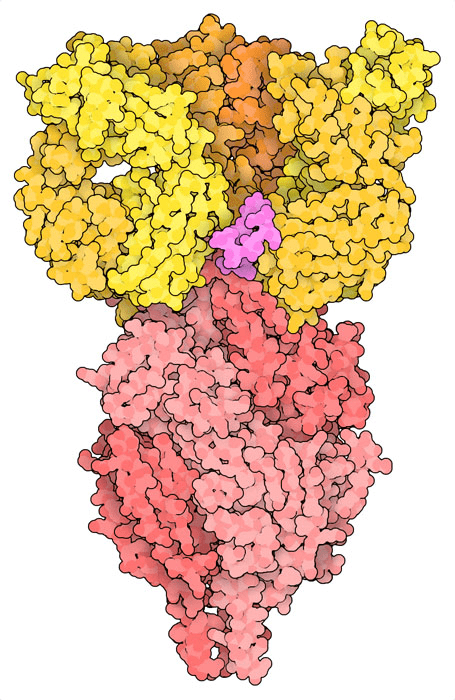
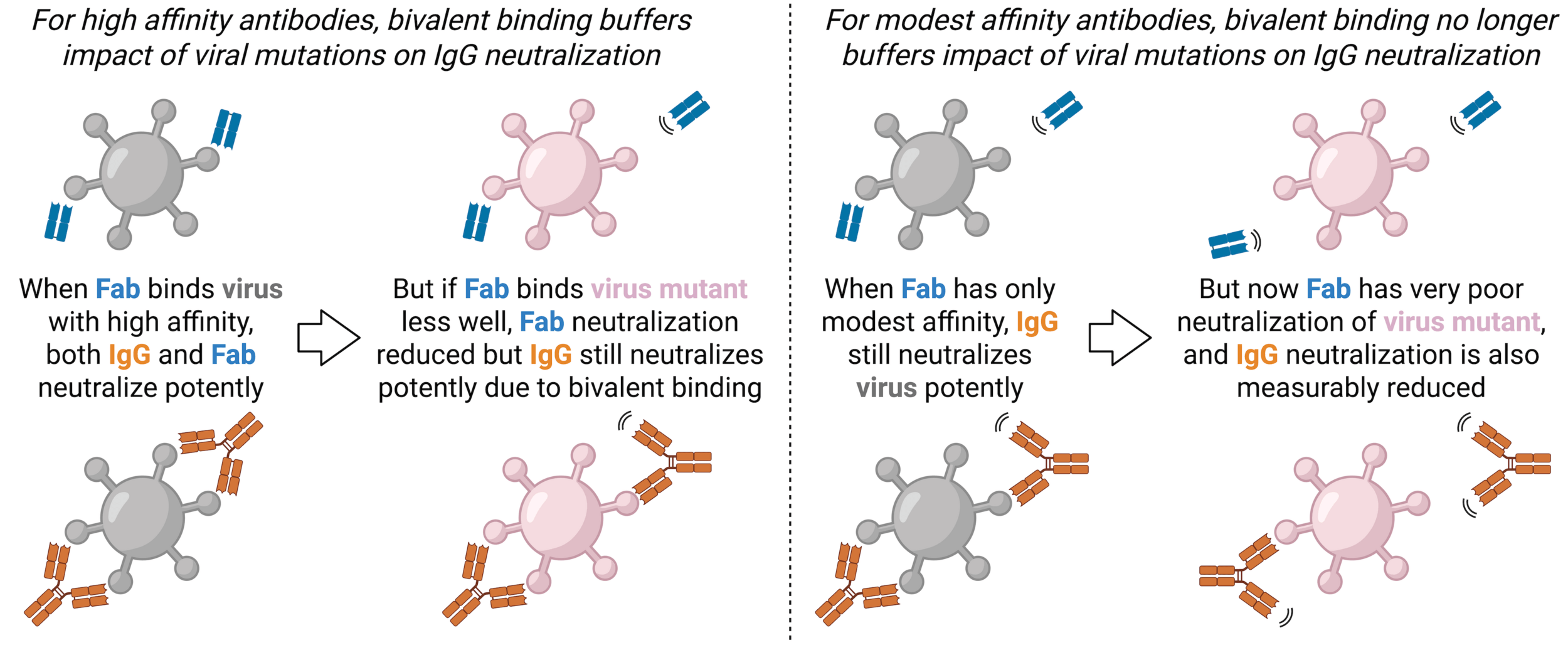
The antibody nirsevimab bound to prefusion F at epitope that includes glycan. Note F is a trimer, and structure shows one antibody Fab bound to each monomer.
The antibody nirsevimab bound to prefusion F at epitope that includes glycan.
Although population disease burden is substantial, annual risk to any individual is low, and impossible to prospectively identify who will become severely ill at any given time.
Monoclonal antibodies are expensive to produce, and repeated dosing is required for sustained protection.
Viruses can evolve to become resistant.
Although population disease burden is substantial, annual risk to any individual is low, and impossible to prospectively identify who will become severely ill at any given time.
Monoclonal antibodies are expensive to produce, and repeated dosing is required for sustained protection.
Viruses can evolve to become resistant.
Severe disease concentrated in an easily identifiable population (infants).
Although population disease burden is substantial, annual risk to any individual is low, and impossible to prospectively identify who will become severely ill at any given time.
Monoclonal antibodies are expensive to produce, and repeated dosing is required for sustained protection.
Viruses can evolve to become resistant.
Severe disease concentrated in an easily identifiable population (infants).
Infants require lower dose (they're smaller), most need protection only for first year, and progress has been made in engineering more potent and long-lived antibodies.
Although population disease burden is substantial, annual risk to any individual is low, and impossible to prospectively identify who will become severely ill at any given time.
Monoclonal antibodies are expensive to produce, and repeated dosing is required for sustained protection.
Viruses can evolve to become resistant.
Severe disease concentrated in an easily identifiable population (infants).
Infants require lower dose (they're smaller), most need protection only for first year, and progress has been made in engineering more potent and long-lived antibodies.
This is a still a concern, and is topic of this talk.
Lower potency than subsequent antibodies.
First approved primarily for prophylaxis of high-risk infants (eg, born prematurely at <36 weeks), with dosing of 15 mg/kg each month for five months.
Due in part to cost, recommendation progressively narrowed: by 2014 only for infants born <29 weeks gestational age or <32 weeks with chronic lung disease
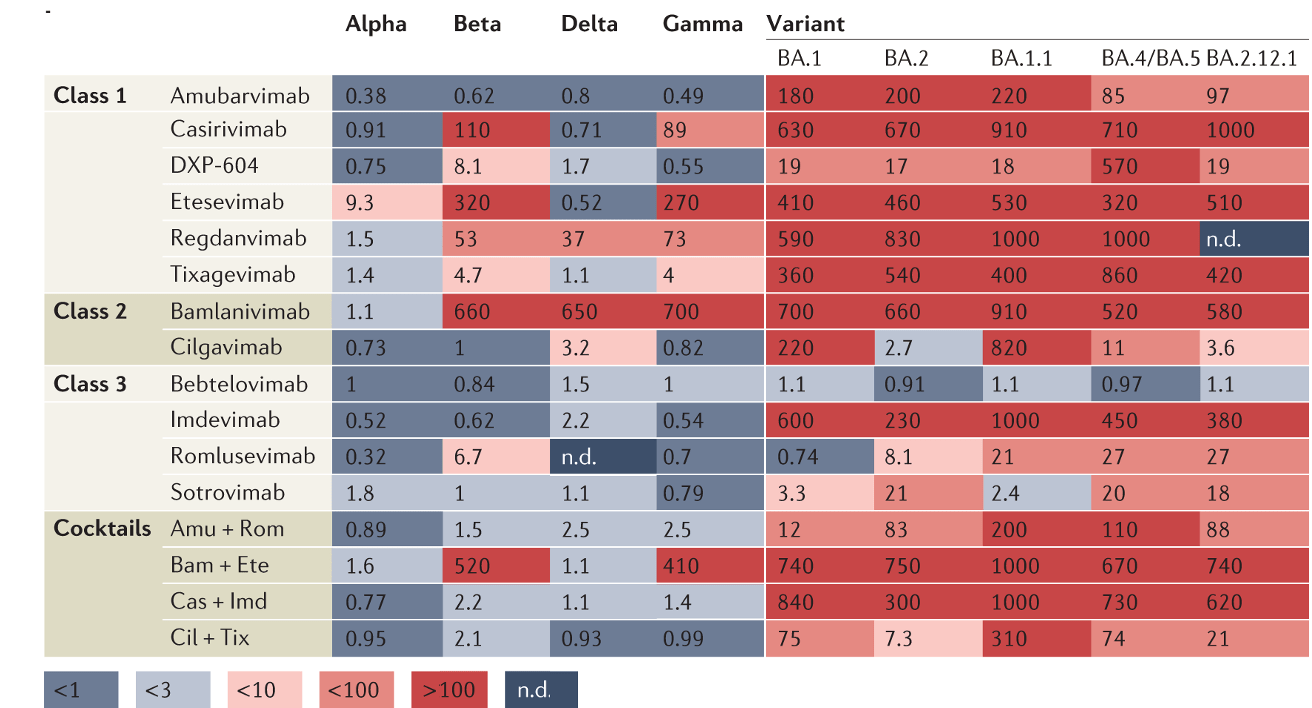
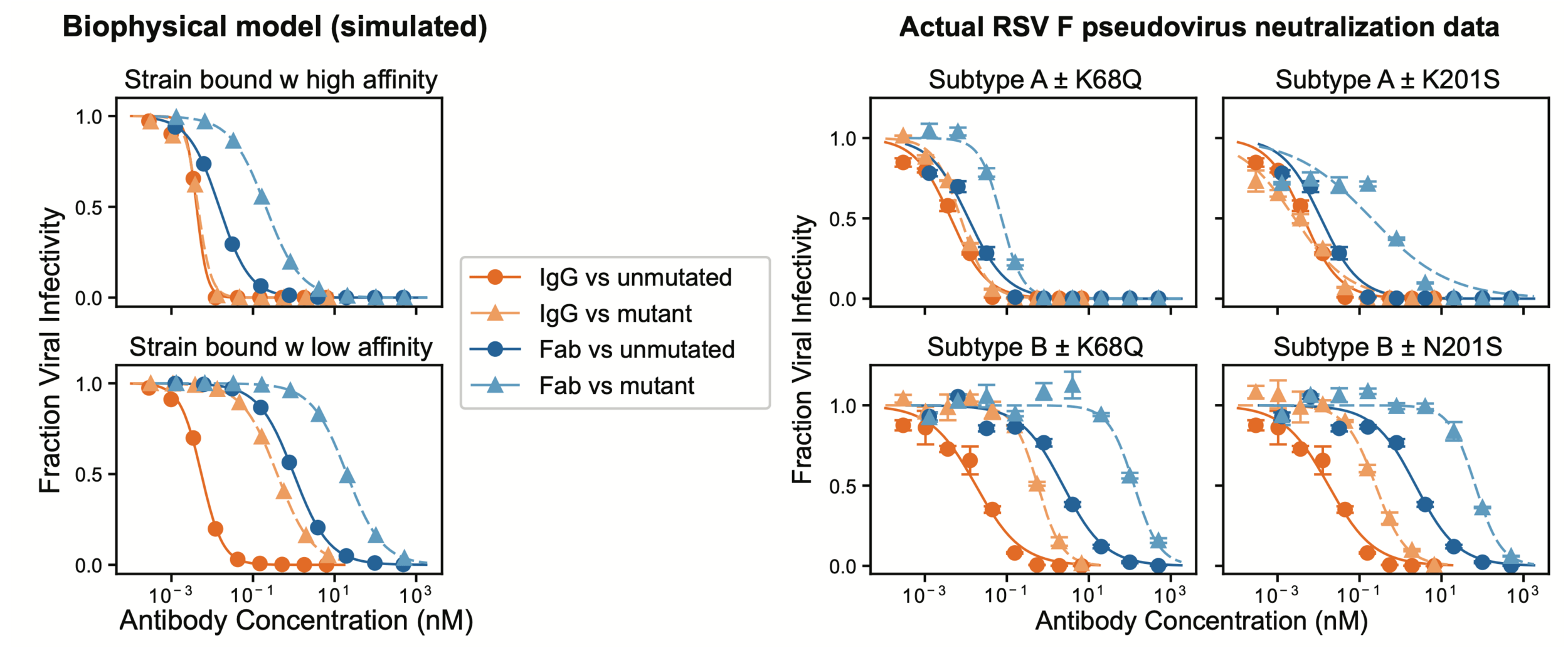
Neutralization curves from Simonich et al (2025)
Developed by Regeneron: much more potent against some strains than palivizumab.
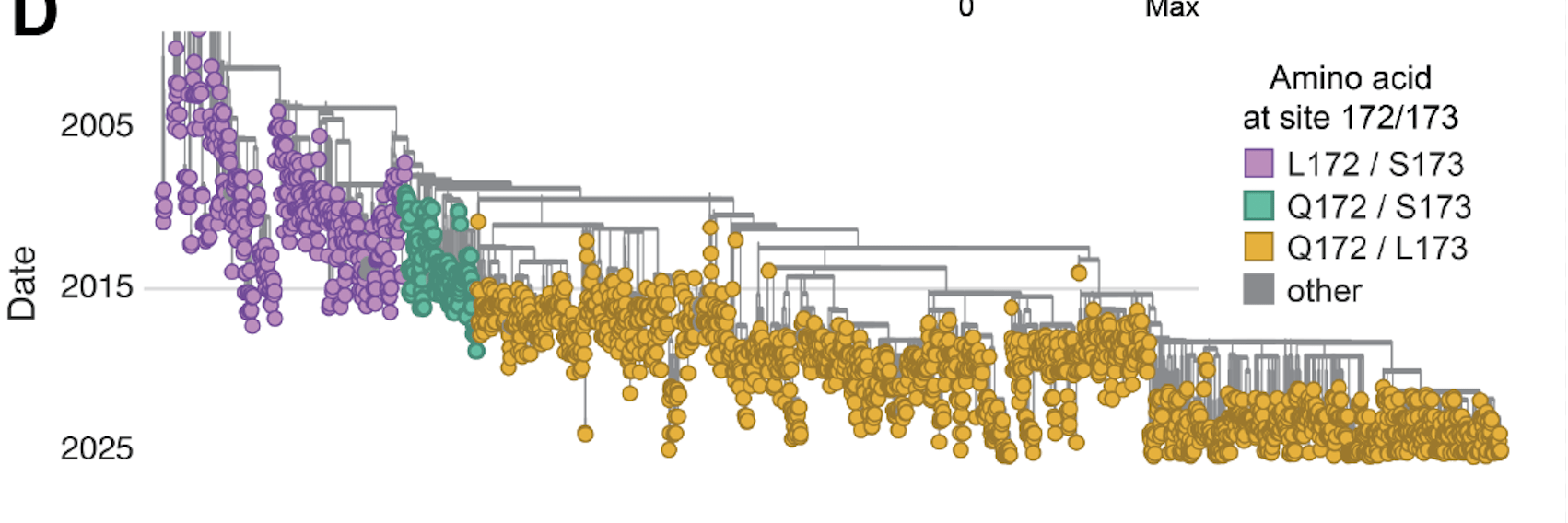
Failed Phase 3 clinical trial from 2015-2017 due to lack of efficacy against subtype B; coincided with evolution of new variants with mutations at F sites 172 and 173.
Neutralization curves from Simonich et al (2025)
Developed by AstraZeneca and Sanofi; much more potent than palivizumab and has extended half life.
Recommended in 2023 for all infants <8 months old entering their first RSV season. Dosing just one injection of 50 mg for infants <5 kg.
~80% effectiveness in preventing RSV hospitalization.
Neutralization curves from Simonich et al (2025)
Developed by Merck, and has high potency and extended half life similar to nirsevimab but targets different region of F.
Similar recommendations for use as nirsevimab, only recently approved (in 2025)
Neutralization curves from Simonich et al (2025)
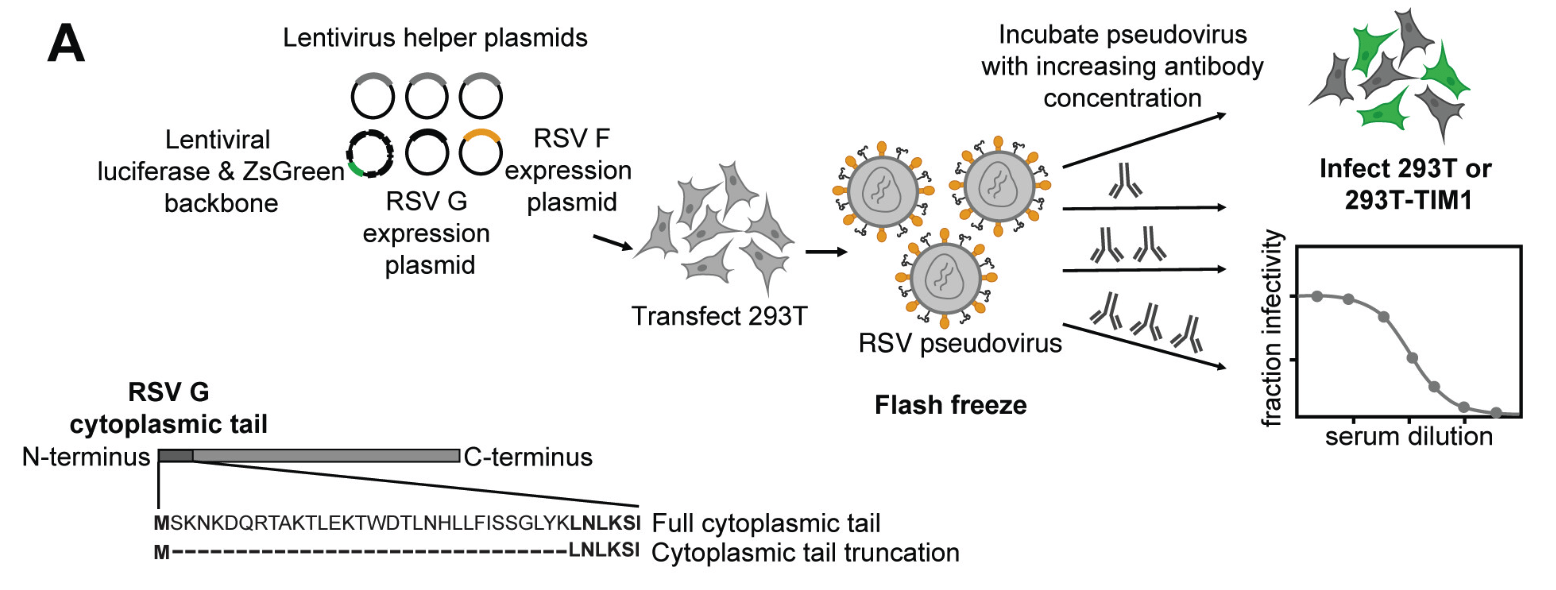
Transfection of RSV F (and G) along with plasmids expressing lentiviral proteins creates pseudotyped viral particles, which can only undergo a single round of cell entry and are not pathogens.
We can synthesize any variant of F and make pseudoviruses for neutralization assays, providing a safe way to study the effects of viral mutations on antibody neutralization.
Resistant strains have been identified in clinical and lab-passaging studies, plotted data from Simonich et al (2025)
Library of pseudoviruses expressing all single amino-acid mutants of RSV F.
Pseudoviruses can only undergo single round of cell entry, and so provide safe way to study effect of F mutations.
We can also measure how all mutations affect pseudovirus cell entry in absence of antibody, providing a measure of functional constraint.
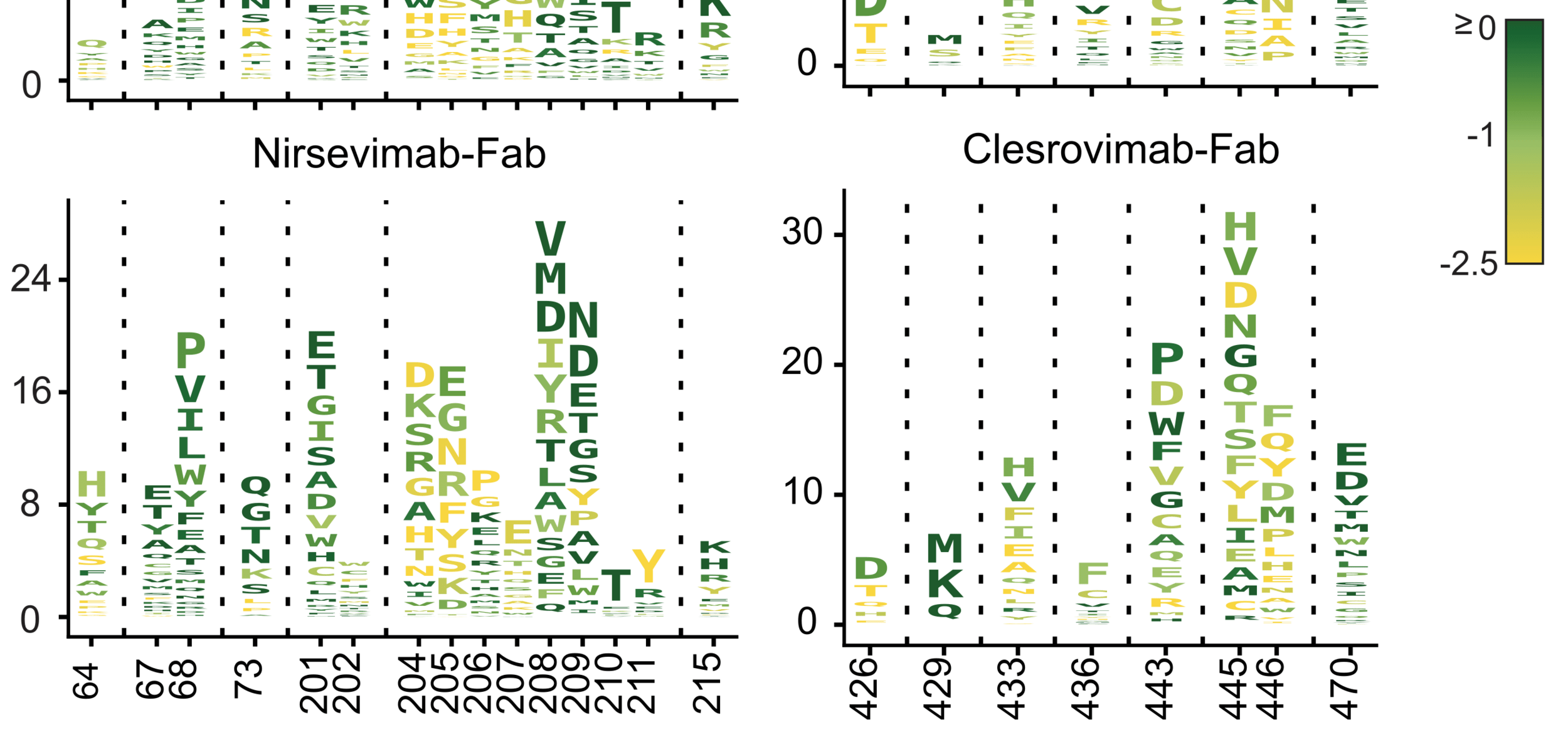
Letter heights indicate reduction in antibody neutralization, color indicates impact on F's cell entry function. These visualizations help quantify how constraint limits escape from different antibodies.
Nirsevimab
Clesrovimab
reduction in neutralization
site
cell entry function
Historically, anti-viral antibodies isolated from infected or vaccinated humans. Those antibodies target whatever regions of a viral protein happen to be immunodominant in human immune response.
In many cases, those regions of a viral protein are also mutationally tolerant and can evolve rapidly. For instance, Jian et al (2022) showed how only 2 of 141 SARS-CoV-2 antibodies isolated from individuals who were exposed to an early SARS-CoV-2 variant still neutralized the viral variants present in 2023. This is because naturally elicited antibodies mostly target mutationally tolerant sites.
Our deep mutational scanning allows us to identify regions of viral proteins that cannot change without disrupting function. If antibodies could be rationally targeted to those epitopes there would be reduced potential for escape, and our deep mutational scanning could quantify this.
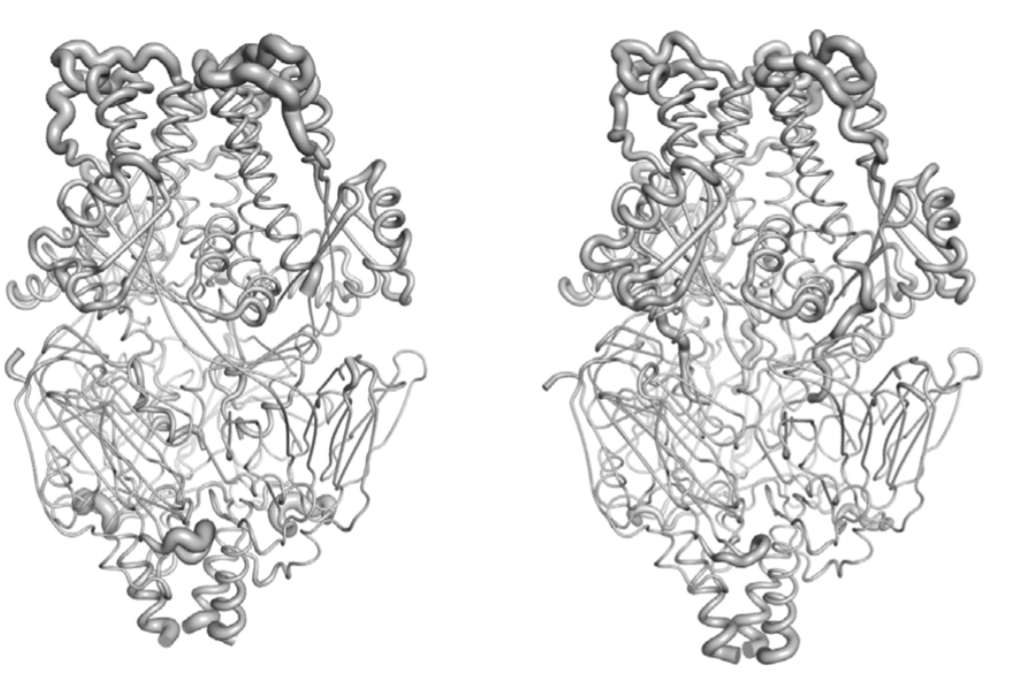
RSV B F structure
RSV A F structure
The structures have a RMSD deviation of only 1.8 angstroms.
plotted data from Simonich et al (2025)
| study | resistance in RSV-A | resistance in RSV-B |
|---|---|---|
| Fourati et al (2025a) | 2/195 = 1% | 23/184 = 13% |
| Fourati et al (2025b) | 0/236 = 0% | 2/24 = 8% |
| Ahani et al (2023) | 0/11 = 0% | 2/14 = 14% |
Rates of resistance to nirsevimab neutralization in RSV breakthrough infections of infants who received nirsevimab.
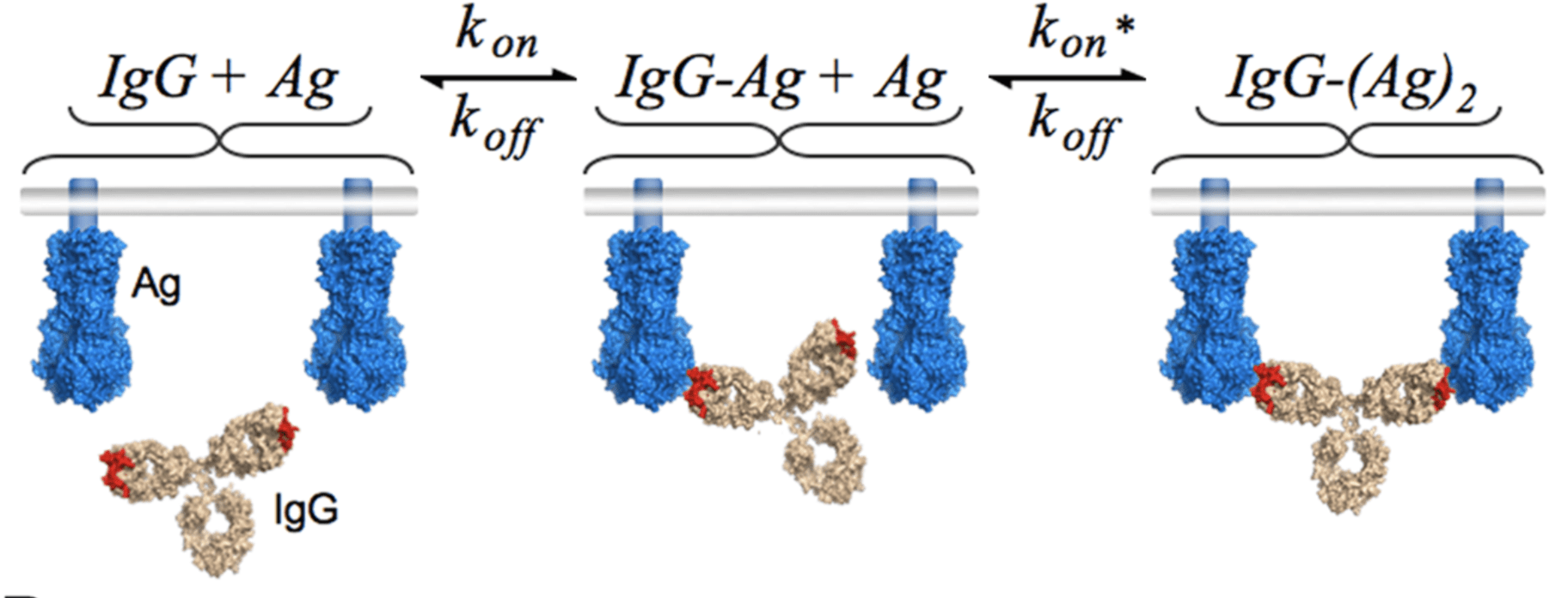
See https://jbloomlab.github.io/IgG-vs-Fab-neutralization/notebook.html for full mathematical model
The only SARS-CoV-2 antibody still approved in USA (VYD222) is a version of an earlier antibody that was re-engineered to target new viral variants (Yuan et al, 2025).
The company producing this has now also reported another generation of engineered antibody (VYD2311) even more potent against new antibodies (Mellis et al, 2026).
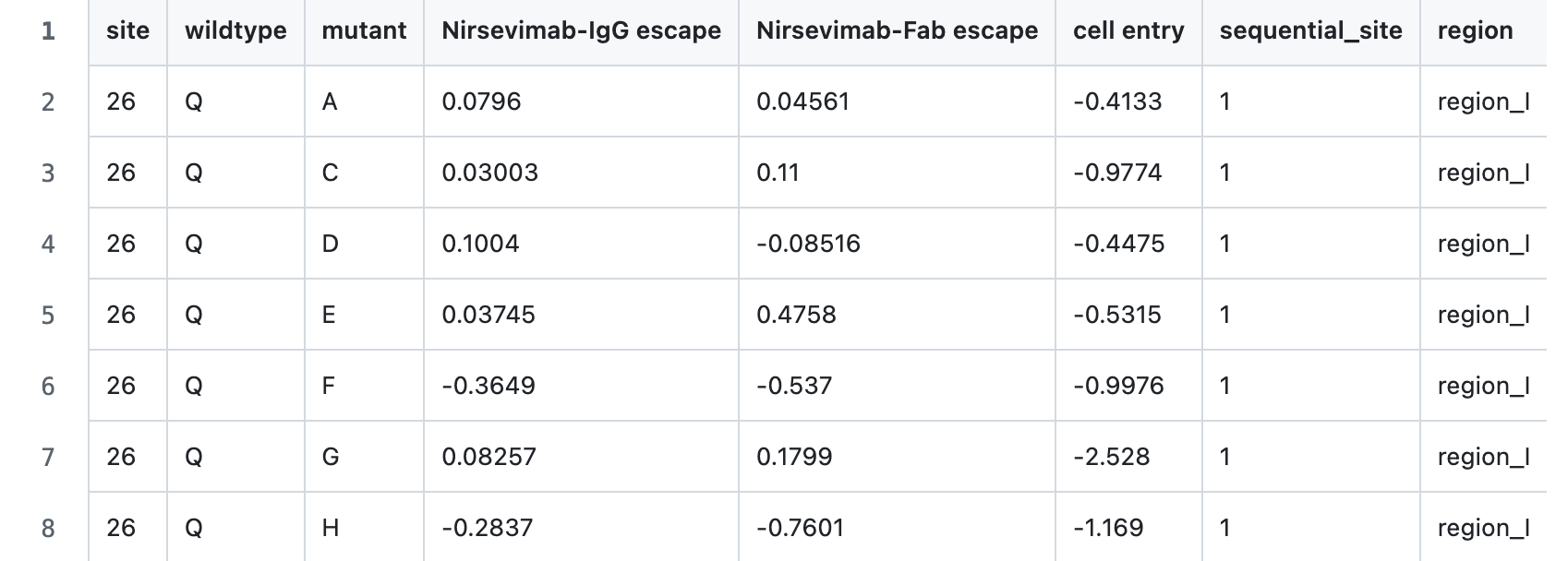
For each mutation relative to a wildtype viral protein sequence, we measure:
We like to look at the data in interactive visualizations like the one here. Elements:
CSVs of numerical data are on GitHub, as per here. These report:
RSV F is an important target, and several major companies (eg, Sanofi, Merck) have antibodies on market, and more are being developed:
SARS-CoV-2 antibodies remain important for vulnerable populations, and two antibodies (VYD222 and VYD2311) are in clinical use or development in USA:
Nipah is a virus of pandemic concern, and antibodies are being developed as countermeasures. RBP is the Nipah receptor-binding protein:
Nipah is a virus of pandemic concern, and antibodies are being developed as countermeasures. F is the Nipah fusion protein:
Avian influenza is considered to be one of the priority pathogens of pandemic concern. There have been 5 influenza pandemics over the last 120 years and most pandemic strains had avian origins. No clinically approved antibody is available for use against influenza viruses.
These slides: https://slides.com/jbloom/bloom-baker-dms-for-ab-design
By Jesse Bloom
How deep mutational scanning of viral proteins might aid do novo design of antibody countermeasures (brainstorming slides for Bloom and Baker labs)




