









Jesse Bloom PRO
Scientist studying evolution of proteins and viruses.
Fred Hutch Cancer Research Center / HHMI
Slides at https://slides.com/jbloom/activ-trace
We studied CoV-229E, a coronavirus that causes common colds and has been circulating in humans since at least the 1960s.
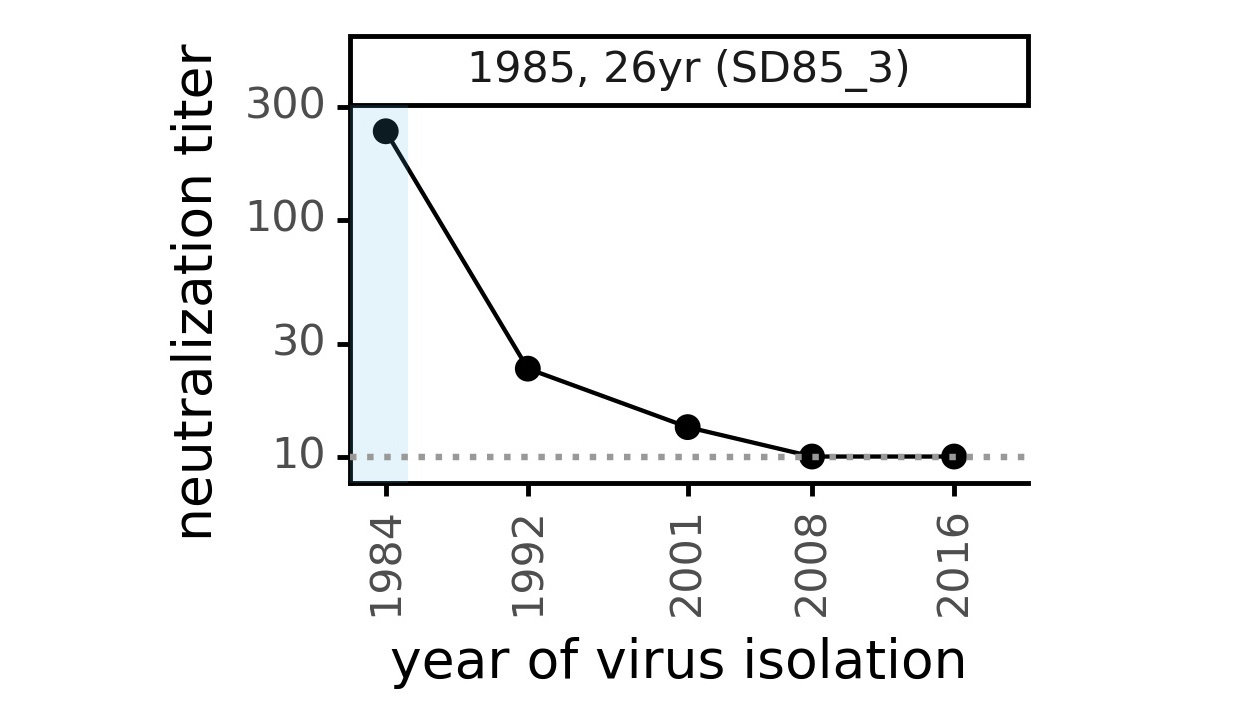
We experimentally generated CoV-229E spikes at ~8 year intervals so we could study them in the lab:
- 1984
- 1992
- 2001
- 2008
- 2016
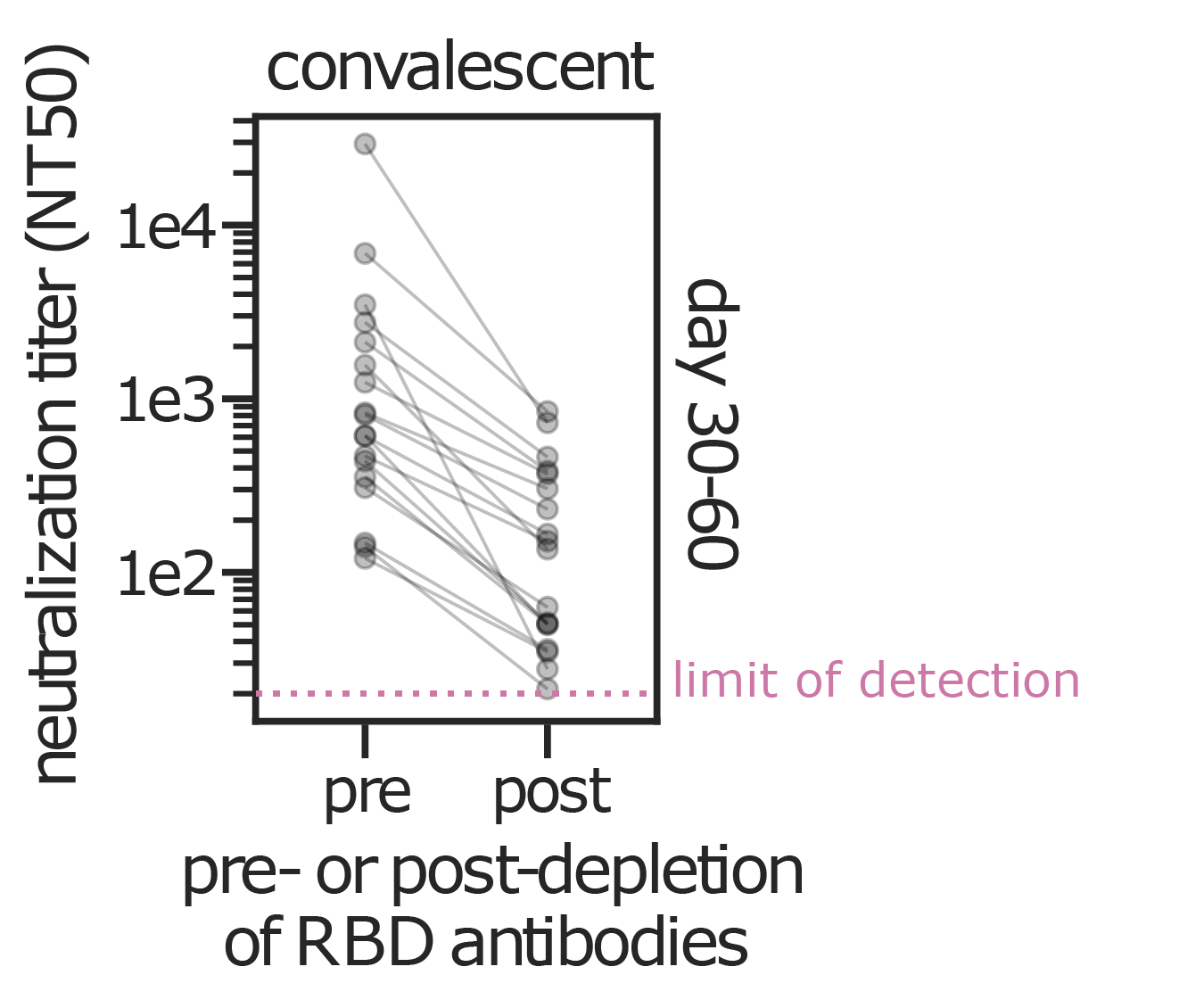
Serum collected in 1985 neutralizes virus with spike from 1984, but less effective against more recent viruses.
We are studying basis of these differences, as ideally vaccines would elicit more evolution-resistant sera as on the right.
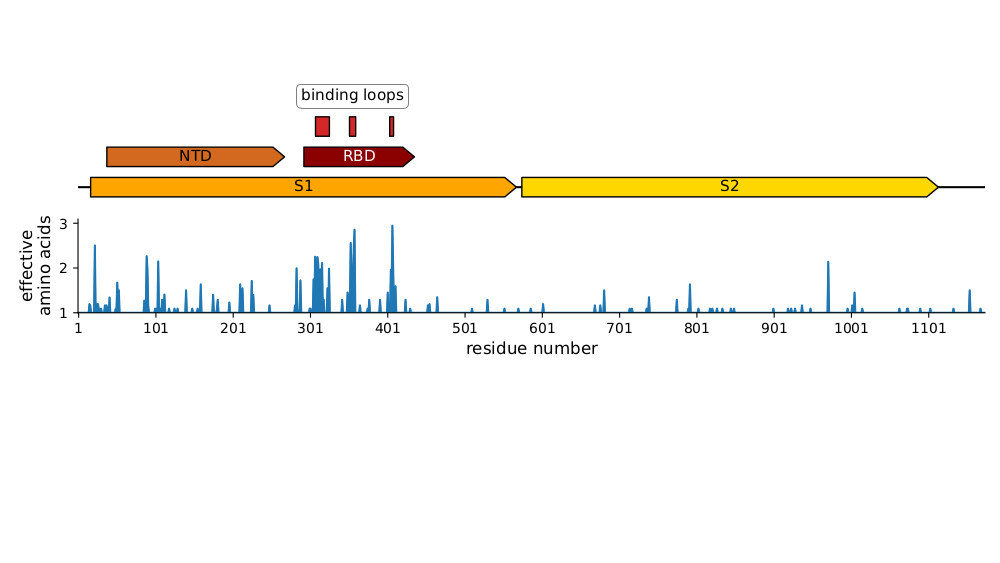
Plot of sequence variability across CoV-229E spike taken from Eguia, ..., Bloom, PLoS Pathogens (2021) . See also Wong, ..., Rini, Nature Communications (2017) and Li, ..., Rini, eLife (2019) for detailed structural studies of evolution in receptor-binding loops.
Impossible to measure true viral fitness in the lab, so we focus on three biochemical phenotypes that contribute to fitness:
1) Does RBD fold properly?
2) Does RBD bind ACE2 with high affinity?
3) Is RBD bound by anti-viral antibodies?
Impossible to measure true viral fitness in the lab, so we focus on three biochemical phenotypes that contribute to fitness:
1) Does RBD fold properly?
2) Does RBD bind ACE2 with high affinity?
3) Is RBD bound by anti-viral antibodies?
Evolutionary pressure is to maintain these two phenotypes...
... while changing this phenotype.
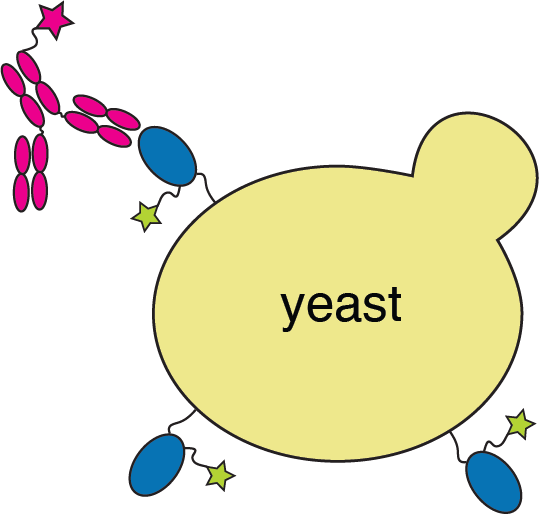
RBD
fluorescent ACE2
yeast
fluorescent tag on RBD
Importantly, we use ACE2 titrations to measure true affinities, not just relative FACS binding signal; see here for details.
Click here for details on how library is made.
RBD
fluorescently labeled antibody
yeast
fluorescent tag on RBD
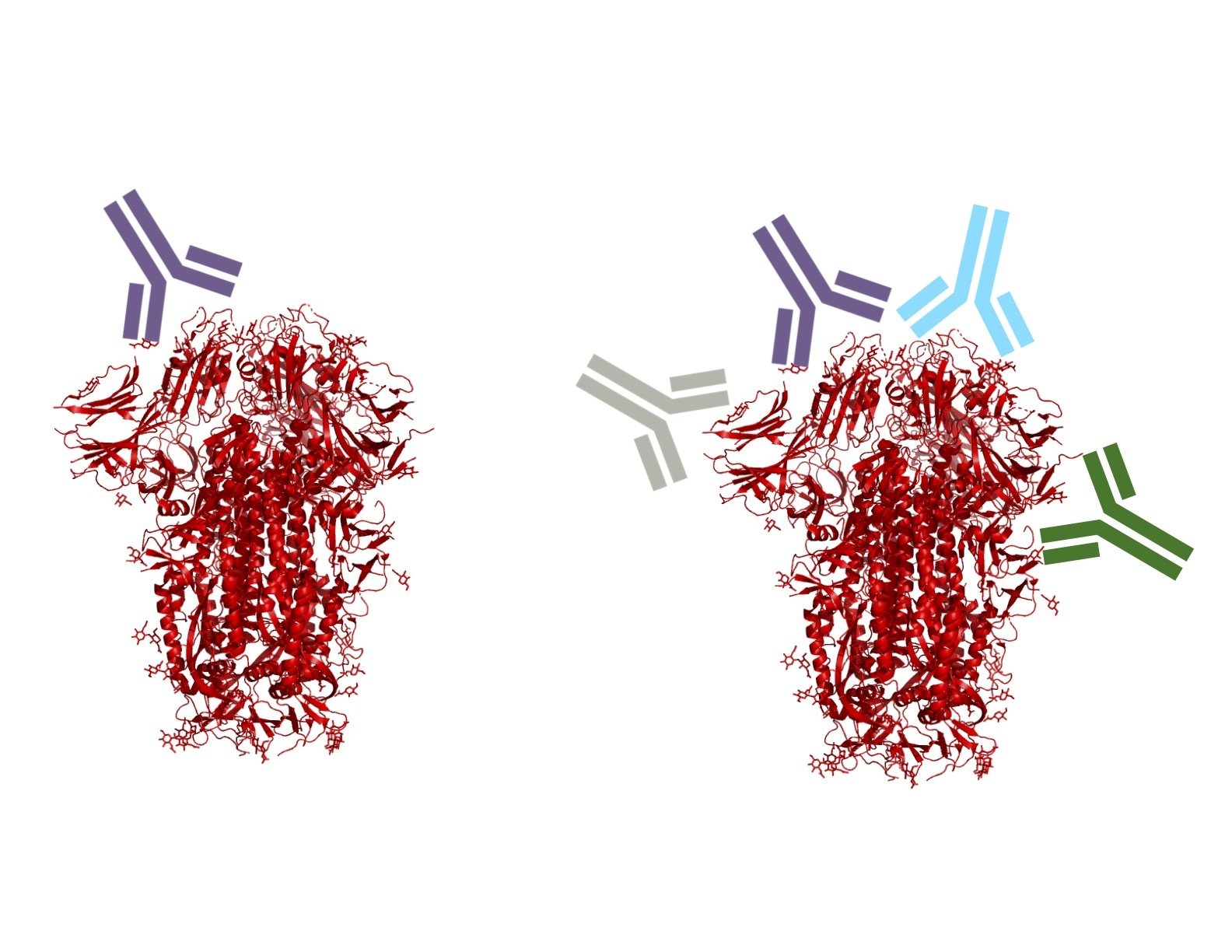
Monoclonal antibodies bind one epitope, so can usually be escaped by single mutation
Polyclonal antibodies can bind many epitopes, so often more resistant to escape
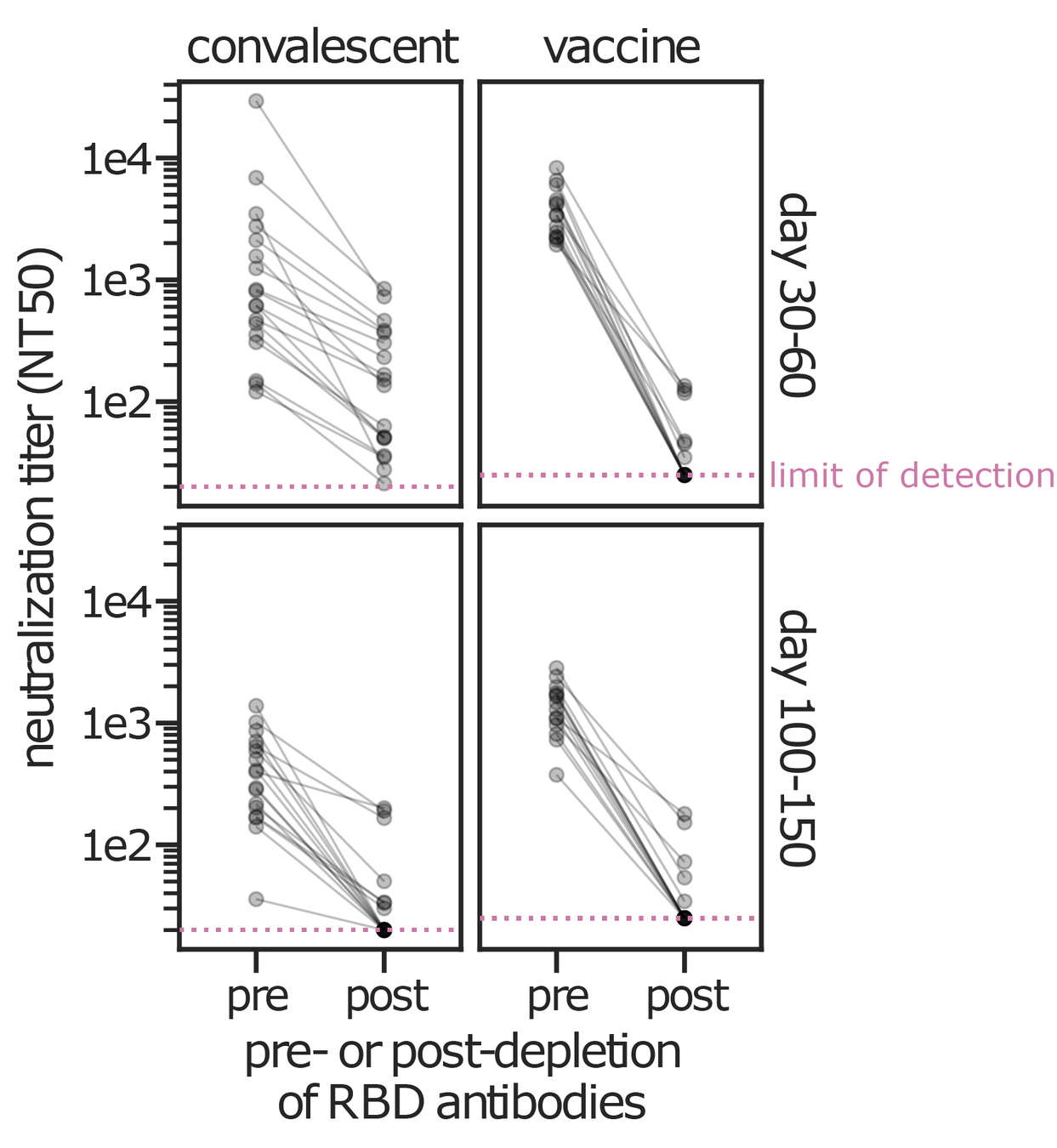
Data from Greaney et al (2021a, 2021b) using lentiviral pseudotypes on ACE2-overexpressing cells. Similar results seen by Piccoli et al (2020). Neutralizing antibodies can target other spike regions such as NTD (e.g., McCallum et al, 2021).
Data from Greaney et al (2021a, 2021b) using lentiviral pseudotypes on ACE2-overexpressing cells. Similar results seen by Piccoli et al (2020). Neutralizing antibodies can target other spike regions such as NTD (e.g., McCallum et al, 2021).
Data from Greaney et al (2021a, 2021b) using lentiviral pseudotypes on ACE2-overexpressing cells. Similar results seen by Piccoli et al (2020). Neutralizing antibodies can target other spike regions such as NTD (e.g., McCallum et al, 2021).
We use the Barnes antibody classification scheme from Barnes et al Nature (2020). The extension of these antibody classes to escape mapping is described in Greaney et al (2021), and escape maps are available here. Class 1, 2, and 3 antibodies are often potently neutralizing, while class 4 antibodies are often less neutralizing (see: Piccoli et al (2020), Dejnirattisai et al (2021), Liu et al (2020), Zost, et al (2020)).
Crowe lab (Vanderbilt): James Crowe, Seth Zost, Pavlo Gilchuk
Chu lab (Univ Wash): Helen Chu, Caitlin Wolf
Veesler lab (Univ Wash): David Veesler, Alexandra Walls, Ale Tortorici
King lab (Univ Wash): Neil King, Dan Ellis
Li lab (Brigham & Women's): Jonathan Li, Manish Choudhary
Whelan lab (Wash U)
Boeckh lab (Fred Hutch): Terry Stevens-Ayers
Alex Greninger (Univ Wash)
Janet Englund (Seattle Children's)
Adam Dingens
Will Hannon
Amin Addetia
Keara Malone
Tyler Starr
Allie Greaney
Rachel Eguia
Bloom lab (Fred Hutch)
Sarah Hilton
Kate Crawford
Andrea Loes
By Jesse Bloom
Interpreting the evolution of SARS-CoV-2



