












Jesse Bloom PRO
Scientist studying evolution of proteins and viruses.
Fred Hutch Cancer Center / HHMI
These slides at https://slides.com/jbloom/embo-workshop
viral membrane
cell membrane
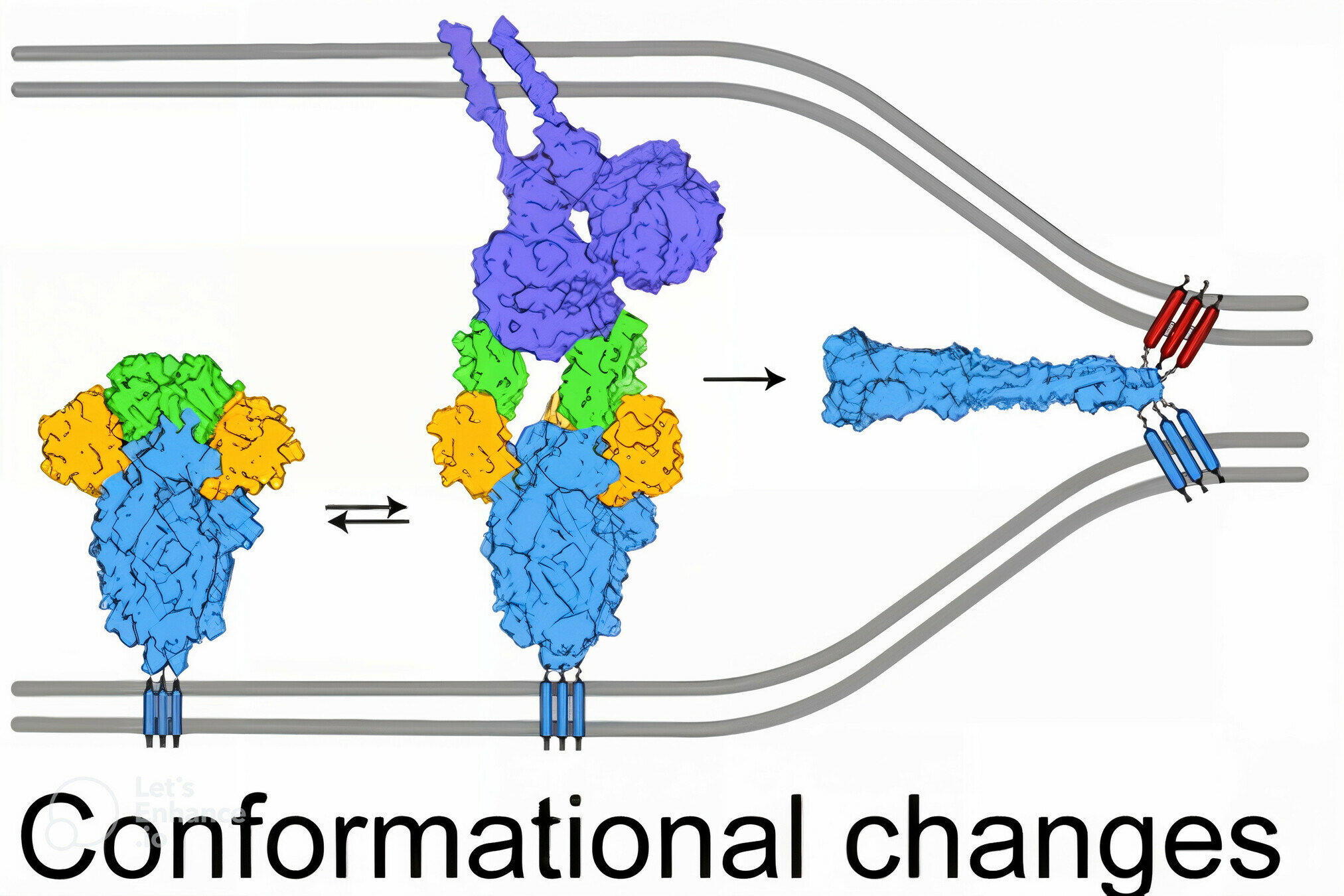
spike
spike conformational change
Image adapted from here
ACE2
antibody
Image adapted from here
RBD
fluorescent ACE2
yeast
fluorescent tag on RBD
cell sorting
RBD
fluorescently labeled antibody
yeast
fluorescent tag on RBD
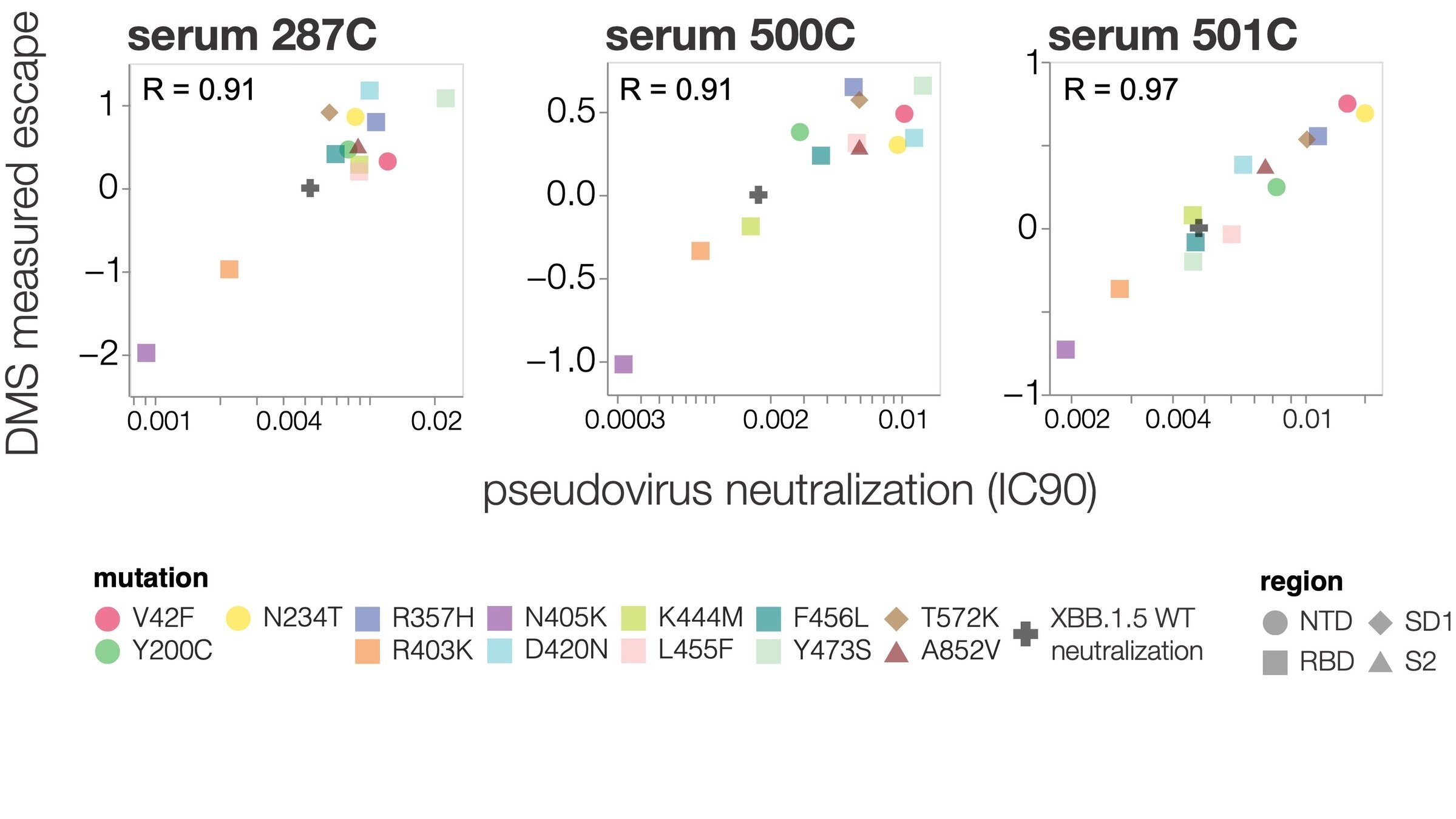
site in RBD
antibody escape
484
cell sorting
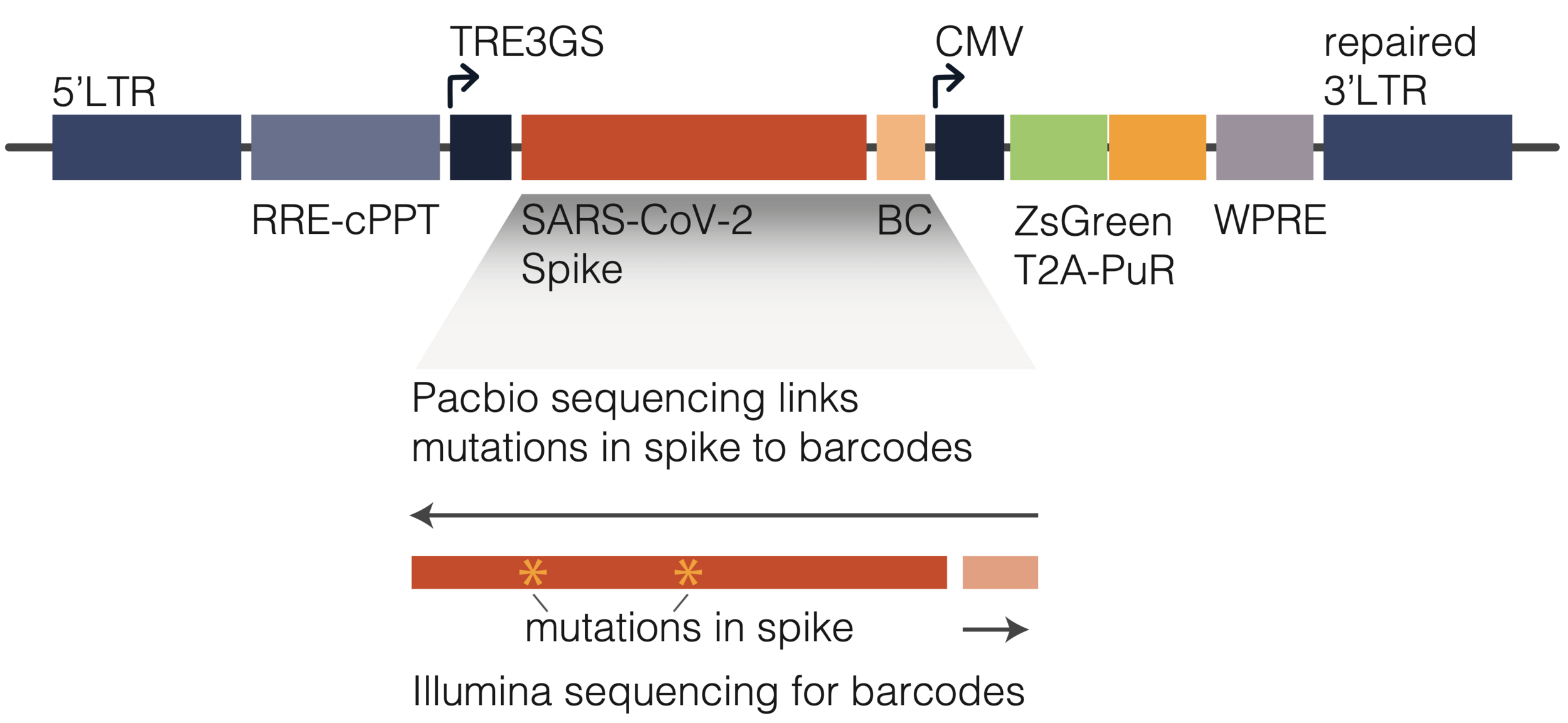
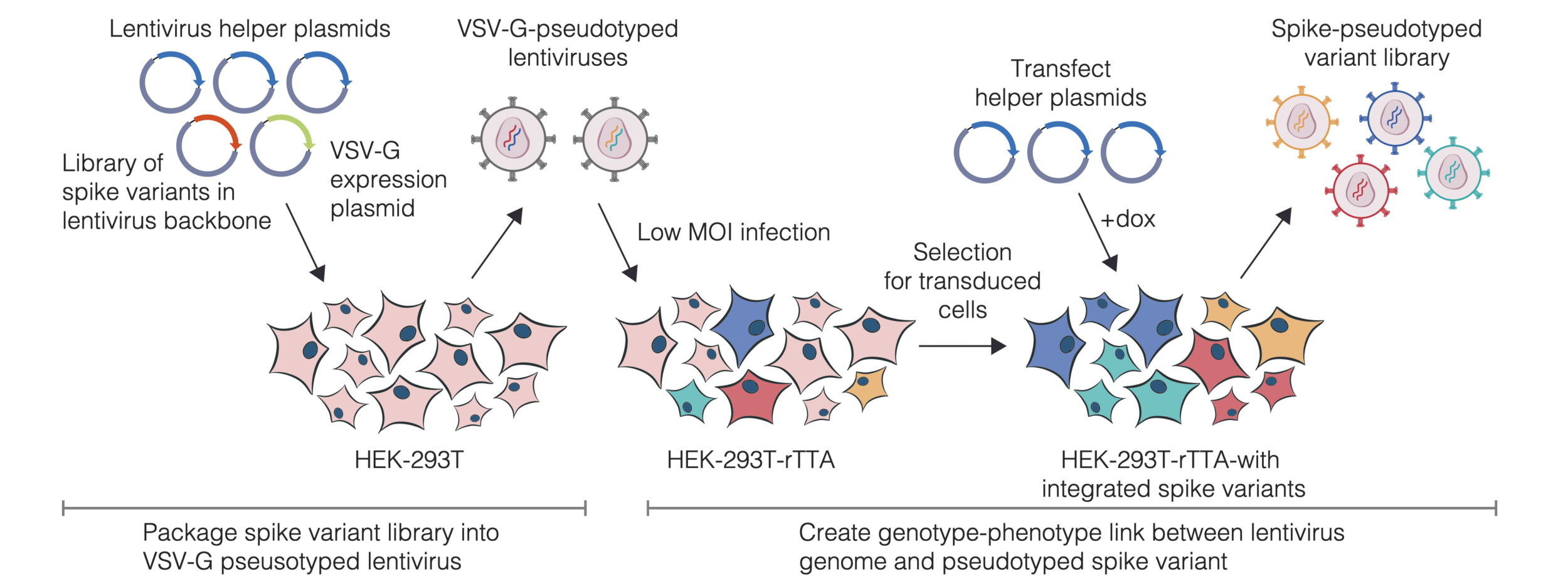
actual SARS-CoV-2 virion: pathogen capable of spread in humans
pseudotyped lentiviral particle: not a pathogen, cannot spread in humans
actual SARS-CoV-2 virion: pathogen capable of spread in humans
pseudotyped lentiviral particle: not a pathogen, cannot spread in humans
With Trevor Bedford & Ben Murrell
With Trevor Bedford & Ben Murrell
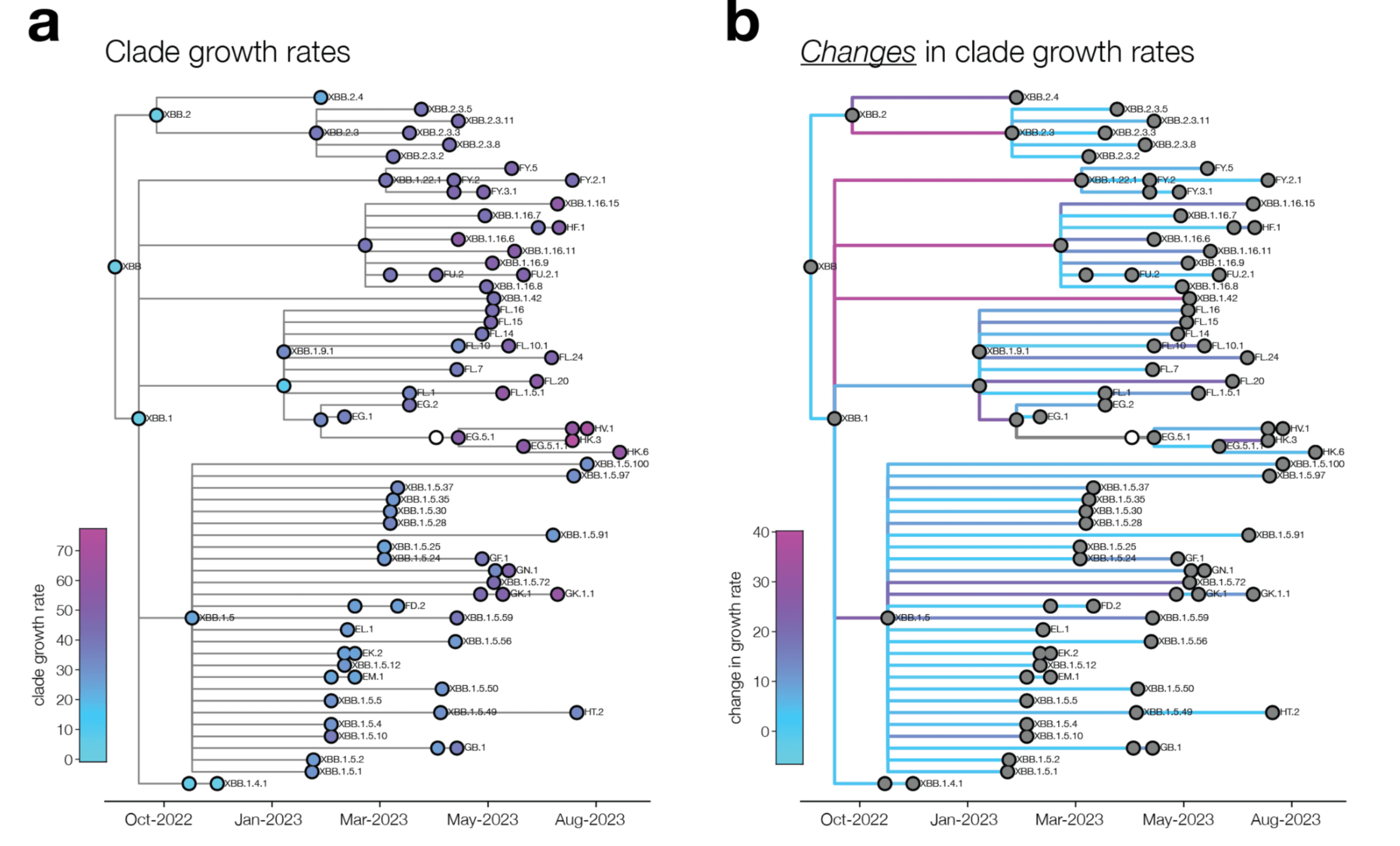
change in clade growth
clade growth
We can explain ~55% of the variance in growth of different clades, with largest fraction of variance uniquely explained by sera escape.
(L122Q, A160T, T199I)
(L122Q, P162Q, T199I)
(L122Q, A160T, T199I)
(L122Q, P162Q, T199I)
For human endemic (SARS-CoV-2) and potential pandemic (H5N1) viruses, we can safely measure how mutations to entry proteins affect key molecular phenotypes.
For SARS-CoV-2, these measurements can help predict success of variants in humans.
For H5N1, these measurements can help inform surveillance of viral evolution.
Bloom lab
Bernadeta Dadonaite
Kate Crawford
Caelan Radford
Tyler Starr
Allie Greaney
Rachel Eguia
William Hannon
Jenny Ahn
Fred Hutch Cancer Center
Trevor Bedford
John Huddleston
University of Washington
Helen Chu and HAARVI cohort
Neil King
David Veesler
Pirbright Institute
Thomas Peacock
University of Pennsylvania
Scott Hensley
Louise Moncla
Jordan Ort
St Jude Children's Hospital
Richard Webby
By Jesse Bloom
Interpreting evolution of SARS-CoV-2 and other viruses



