





Feng Ling PRO
Faculty @ Nankai Physics, studying biophysics, cilia, and flow. Looking for universalities among diversities.
Tap me!
Journey to the Microcosmos (2020)
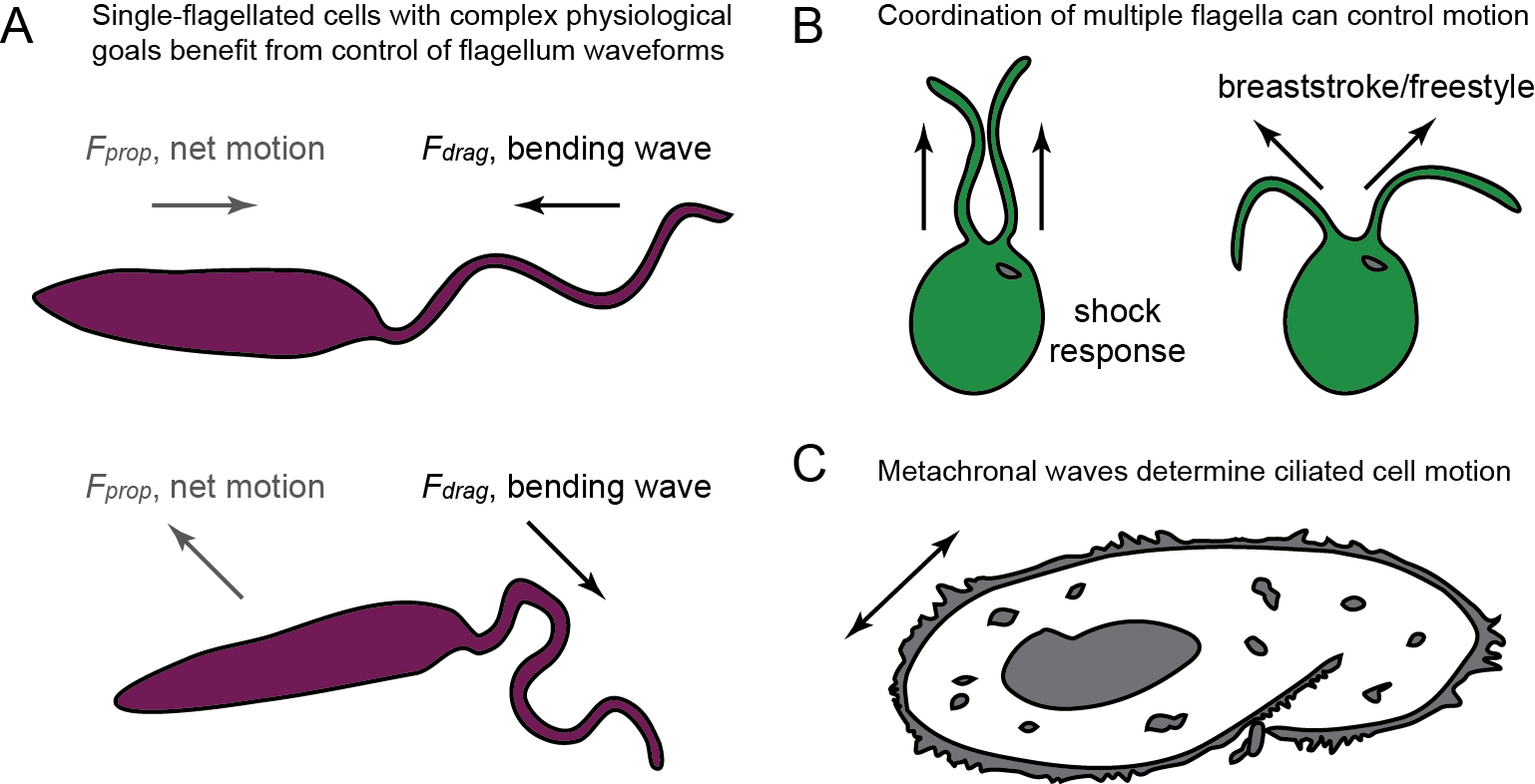
a common navigation strategy is to intersperse a "running/forward" gait with bursts of "tumble/reverse/stop" phase where sensing could occur
this means mono-flagellates need flagellar wave reversals
two model organisms exhibiting wave reversal traits are parasites that separately cause sleeping sickness and Leishmaniasis.
Tip-to-base (T2B) "forward" waves
Base-to-tip (B2T) "reversed" waves
Euglenozoa
Kinetoplastida
Trypanosomatida
T. Brucei
L. Mexicana
metachronal waves
Not all flagellar waves reverse
spermatozoa typically swim with base-to-tip (B2T) waves in helical trajectories
multi-flagellated cells and ciliated microorganisms can change locomotion strategies by altering cilia/flagella synchronization
Lin and Nicastro (2018)
Lin and Nicastro (2018)
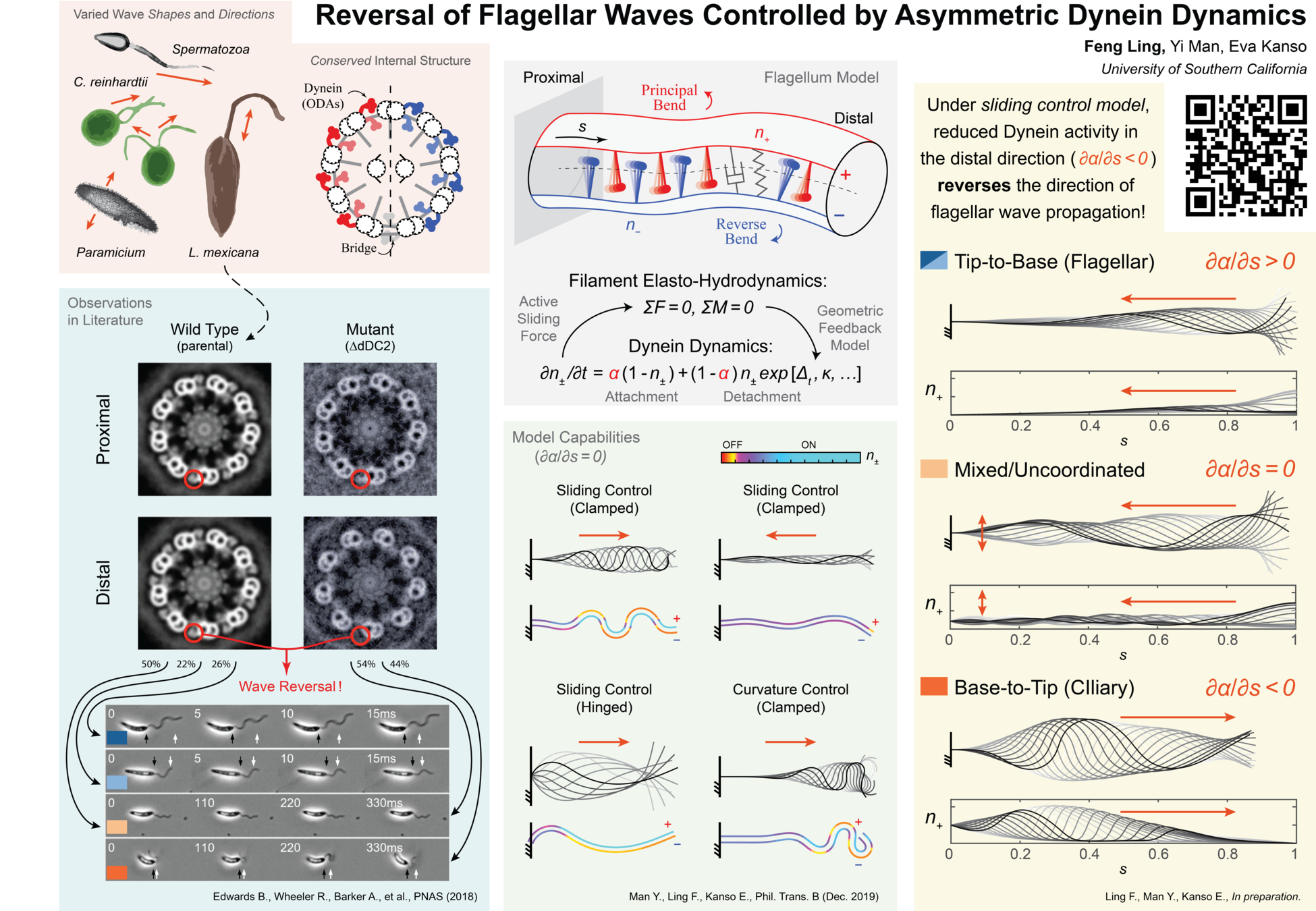
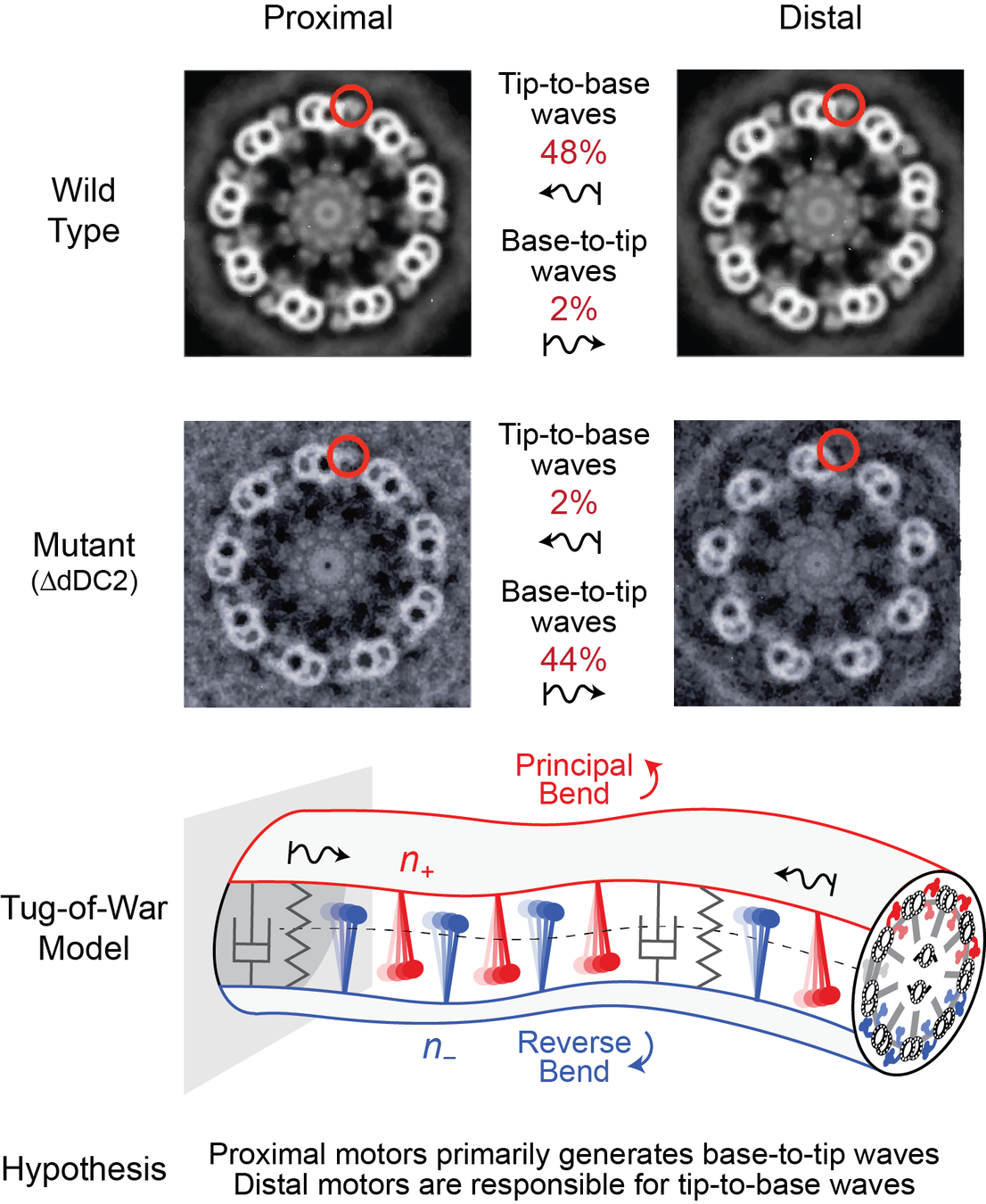
experiments on genetic mutants link wave reversal to asymmetry in dynein motors
specifically, wild type organism typically exhibit tip-to-base (T2B) waves, while mutants missing dynein arms near the tip favor base-to-tip (B2T) waves*
can/do wild type organisms use this asymmetry to control wave direction?
Edwards et al., PNAS (2018)
Tip-to-Base
Mixed/Static
Base-to-Tip
Intermittent T2B
26%
22%
50%
2%
1%
1%
54%
44%
Tap Me !
Edwards et al., PNAS (2018)
model a flagellum as two micro-filaments coupled elastically and via molecular motor dynamics*
each filament (\(\,+\) and \(−\) ) has a population of motors that bend the centerline in opposite directions (\(\,n_+\) and \(n_-\))
Motors attach under a fixed rate but detach due to both natural transitions & geometric feedback
binding/unbinding (attach/detach) of dynein \(\rightarrow\) inter-doublet sliding
sliding force couple + elastic resistance \(\rightarrow\) active bending moment
feedback could arise via sliding control, curvature control, or "geometric clutch"
Qiu et al., Nature Struct. Mol. Bio (2012)
Sartori et al., eLife (2016)
flagellum length
diameter
bending rigidity
drag coefficient
\(L~~~~~~~~~~~\sim50\,[\mu m]\)
\(d~~~~~~~~~~\sim200\,[nm]\)
\(B~~~~~~\sim1\,[nN\,\mu m^2]\)
\(\xi_\perp~~~~ 10^{-3}\sim1\,[Pa\,s]\)
doublet link modulus
total dynein density
\(K~~~~\sim 2\,[\mu N\,\mu m^{-2}]\)
\(\rho~~~~~~~~\lesssim 400\,[\mu m^{-1}]\)
dynein stall force
detachment force
null-force speed
reaction time
\(f_o~~~~~~~~~~~~1\sim5\,[pN]\)
\(f_c~~~~~~0.5\sim2.5\,[pN]\)
\(v_o~~~~~~~5\sim7\,[\mu m\,s^{-1}]\)
\(\tau_o~~~~~~~~~~~\sim50\,[ms]\)
sperm number
\(\text{Sp}^4\;=~\dfrac{\xi_\perp L/\tau_o}{B/L^3}\)
passive moment
duty ratio
char. force
relative disp.
active moment
\(~\mu_a~~=~\dfrac{\rho\,df_o}{B/L^2}\)
\(~\mu~~~~=~\dfrac{Kd^2}{B/L^2}\)
\(~\alpha~~~~=~~~\pi_o\tau_o\)
\(~f^*~~\,=~~~f_o/f_c\)
\(~\zeta~~~\;\,=~~~d/v_o\tau_o\)
dimensional
attachment rate
motor
asymm.
\(f(\alpha)\)
active moment
\(\mu_a\)
sperm number
\(\text{Sp}\)
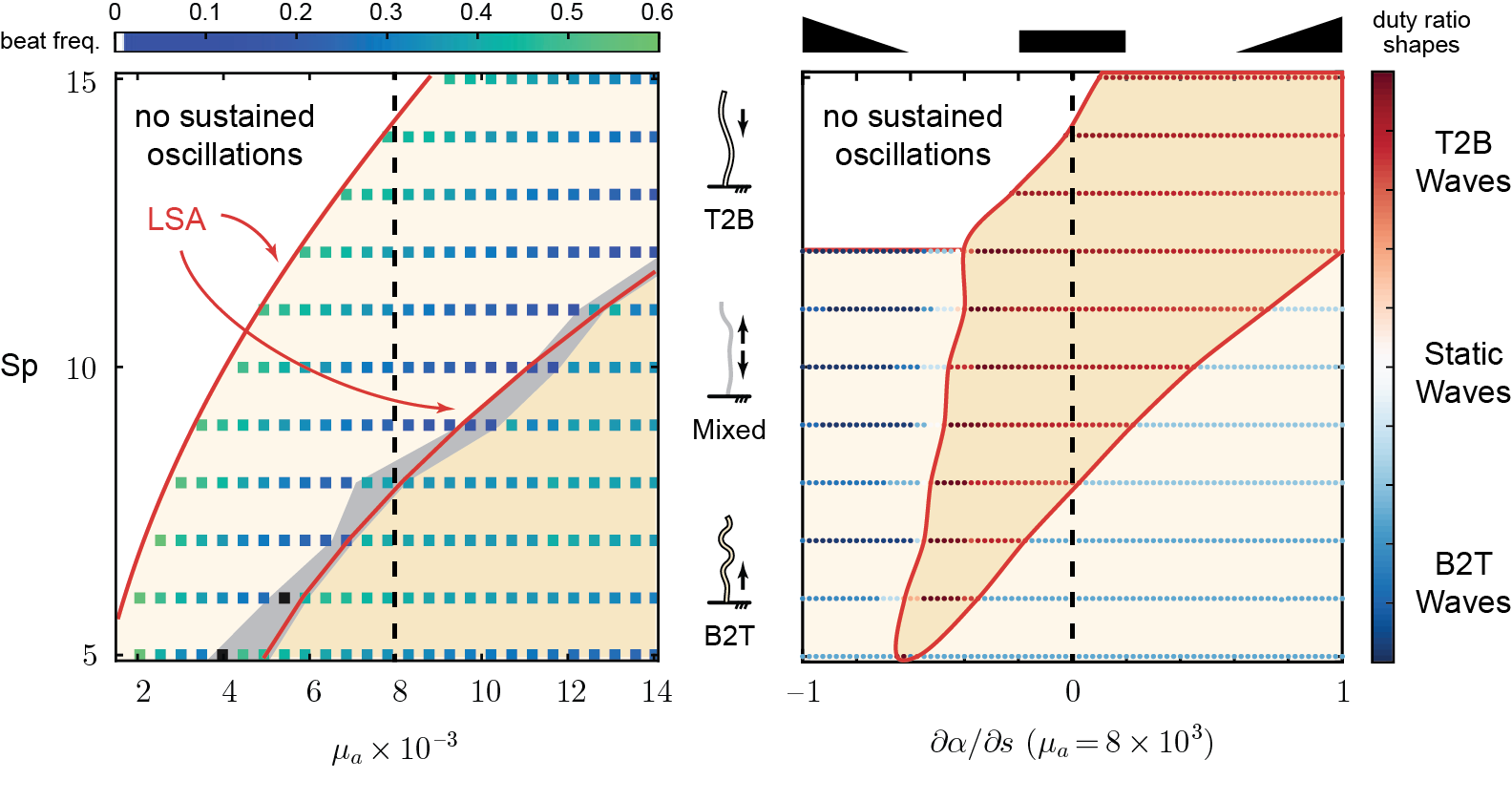
fixing all parameters except \(\text{Sp}\) and dimensionless activity \(\mu_a\), we see that sliding control give rise to
sperm number
active moment
If we fix \(\mu_a\) but vary the attachment duty ratio \(\alpha\) linearly as a function of arclength \(s\)
\(\text{Sp}=10\)
\(\text{Sp}=10\)
\(\text{Sp}=10\)
sperm number
motor asymm.
\(n_+\)
\(n_-\)
\(n_+\)
\(n_-\)
\(n_+\)
\(n_-\)
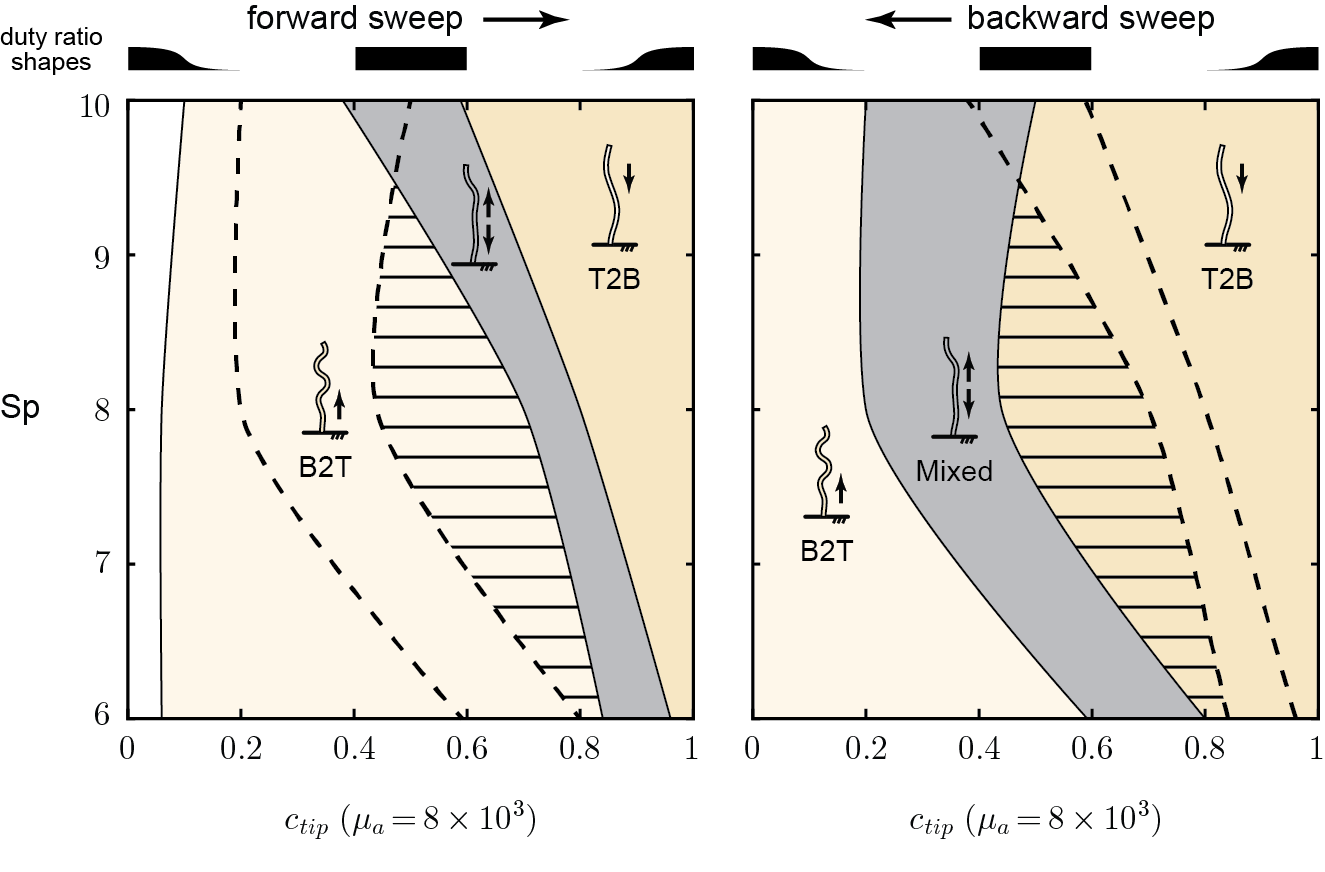
previous results suggests that bi-stability zones could exist
if we gradually vary from a base-heavy activity profile (\(\alpha_{prox}(s)\)) to a tip-heavy profile (\(\alpha_{dist}(s)\)) and back, we should observe
B2T waves \(\rightarrow\) mixed waves \(\rightarrow\) T2B waves \(\rightarrow\) mixed waves \(\rightarrow\) B2T waves
proximal dynein / distal dynein
Edwards et al., PNAS (2018)
\(s\)
\(\alpha(s) = (1-c_{tip})\cdot\alpha_{prox}(s)\)
\(+\;c_{tip}\cdot\alpha_{dist}(s)\)
\(\alpha\)
\(s\)
proximal dynein / distal dynein
Edwards et al., PNAS (2018)
translating bump
switch between two groups
\(\alpha\)
\(s\)
\(s\)
\(s\)
B2T waves are possible even when activity concentrates near the tip! And the opposite is also true!
\(\text{Sp}=8\)
\(\mu_a=8\times 10^3\)
\(n_+\)
\(n_-\)
\(\text{Sp}=8\)
\(\mu_a=8\times 10^3\)
\(\mathbf{f}_h\cdot \mathbf{n}\)
\(\mathbf{f}_h\cdot\mathbf{n}\)
\(n_+\)
\(n_-\)
motor activity distributed in the shaded region can support both B2T and T2B waves!
sperm number
motor asymm.
increase in total activity magnitude introduces a T2B to B2T transition under sliding control*
biasing activity at either end leads to wave initiations near that end in general, e.g., base heavy activity \(\rightarrow\) B2T waves
there exists a bi-stable region where same activity pattern can produce different wave propagation directions
comparable to real data, but
difficult to estimate accurate activity parameters and characteristic scales in geometric feedback
other chemical details† and 3D geometrical effects could further complicate this story
results discussed utilize only sliding control
combination of different feedback mechanism might better explain diverse behaviors
effects on propulsive efficiency*
LSA about traveling wave states
B2T waves can be triggered by either a lack of tip motors or more Ca\(^{2+}\)
T2B waves at higher frequency => faster straight swimming
(asymmetric) B2T waves are at larger amplitude but much slower frequency => helical slow swimming
Geometric clutch theories are popular but there are separate evidences on how curvature/sliding control fits wave shapes better
Wave direction change is impossible without changing direction of motor detachment
frequency (slowly) increases with increased activity
do not produce (very) large amplitude waveforms
Base-to-tip wave can be triggered by either a lack of tip activity or an increase in "activity"
Tip-to-base wave produces smaller net forces (slower swimming)
frequency decreases with "activity increase"
Show/prove why direction cannot change under curvature control?
Construct a threshold-based sliding control analytical model assume
\[\theta(s,t) = T(s) \sin(ks-\omega t)\]
\[m_a(s,t) = M(s) \sin(ks-\omega t)\]
\[n_+(s,t) = N(s) \sin(ks-\omega t)\]
\[n_-(s,t) = N(s) \cos(ks-\omega t)\]
2 DiffEQs coupling T,M w/ BCs (linearized elastic filament eqn)
1 Eqn coupling M and N and T (expression of active torque)
1 Eqn coupling N and T (dynamics on motor)
By Feng Ling
APS DFD 2020 Session ID U02.10 https://meetings.aps.org/Meeting/DFD20/Session/U02.10




