













Stefan Sommer
Professor at Department of Computer Science, University of Copenhagen
Stefan Sommer, Rasmus Nielsen, Christy Hipsley, Mads Nielsen
UCPH Data+
Darwin 1859
Until ca. 1980: morphological characters
After ca. 1980: DNA
Corbett-Deitig et al. MBE 37: 160-1614
Questions:
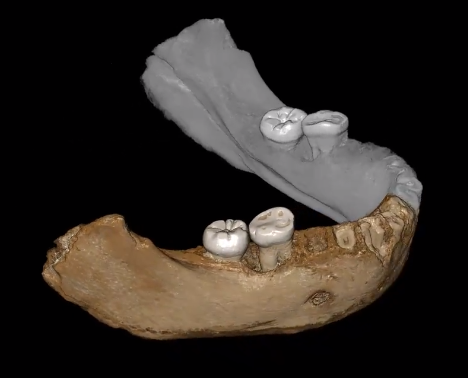
Rules of morphological change
Drivers of morphological change (ecology, historical contingency)
Mechanisms of morphological change (genetic basis)
Current state-of-the-art:
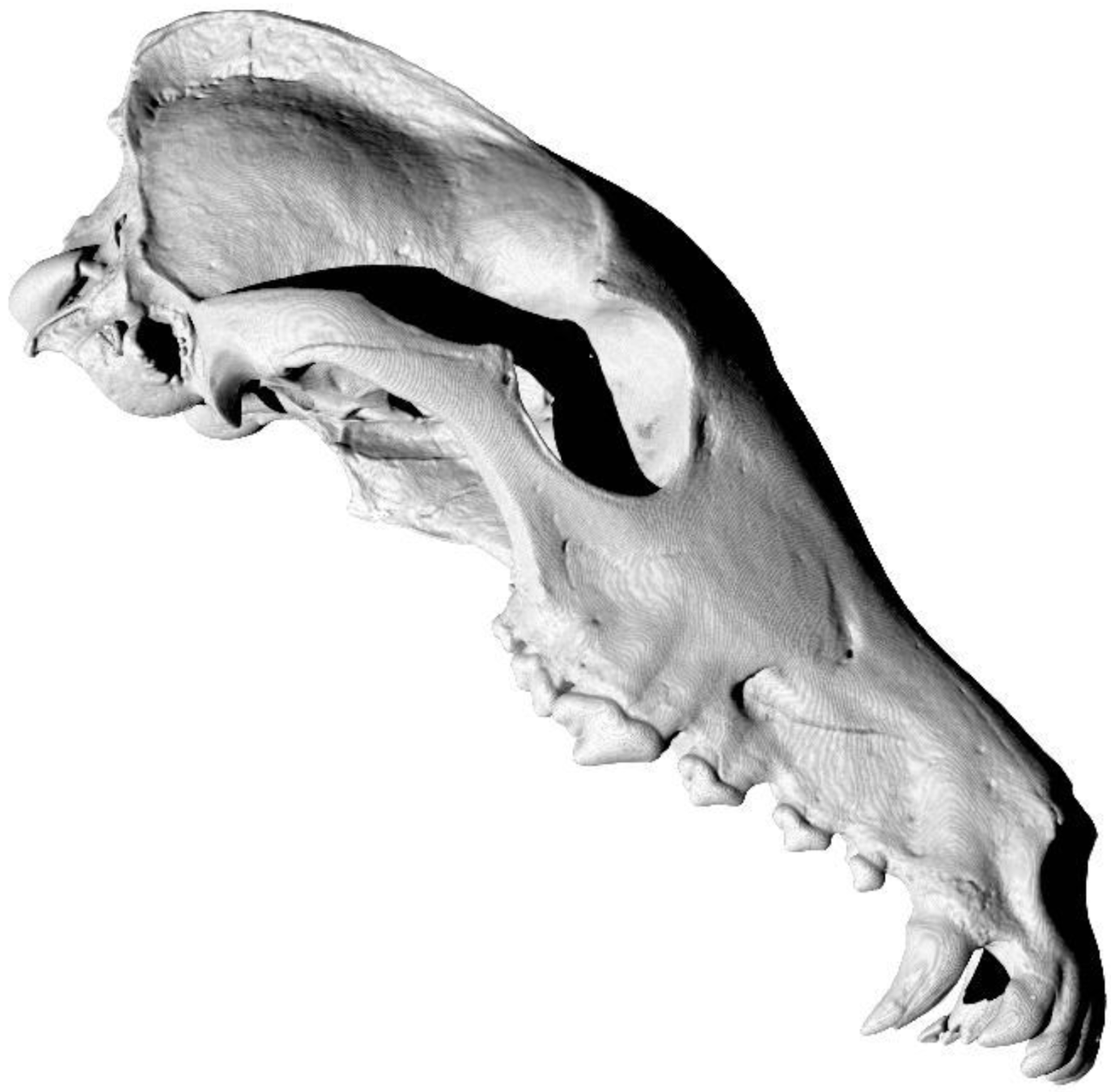
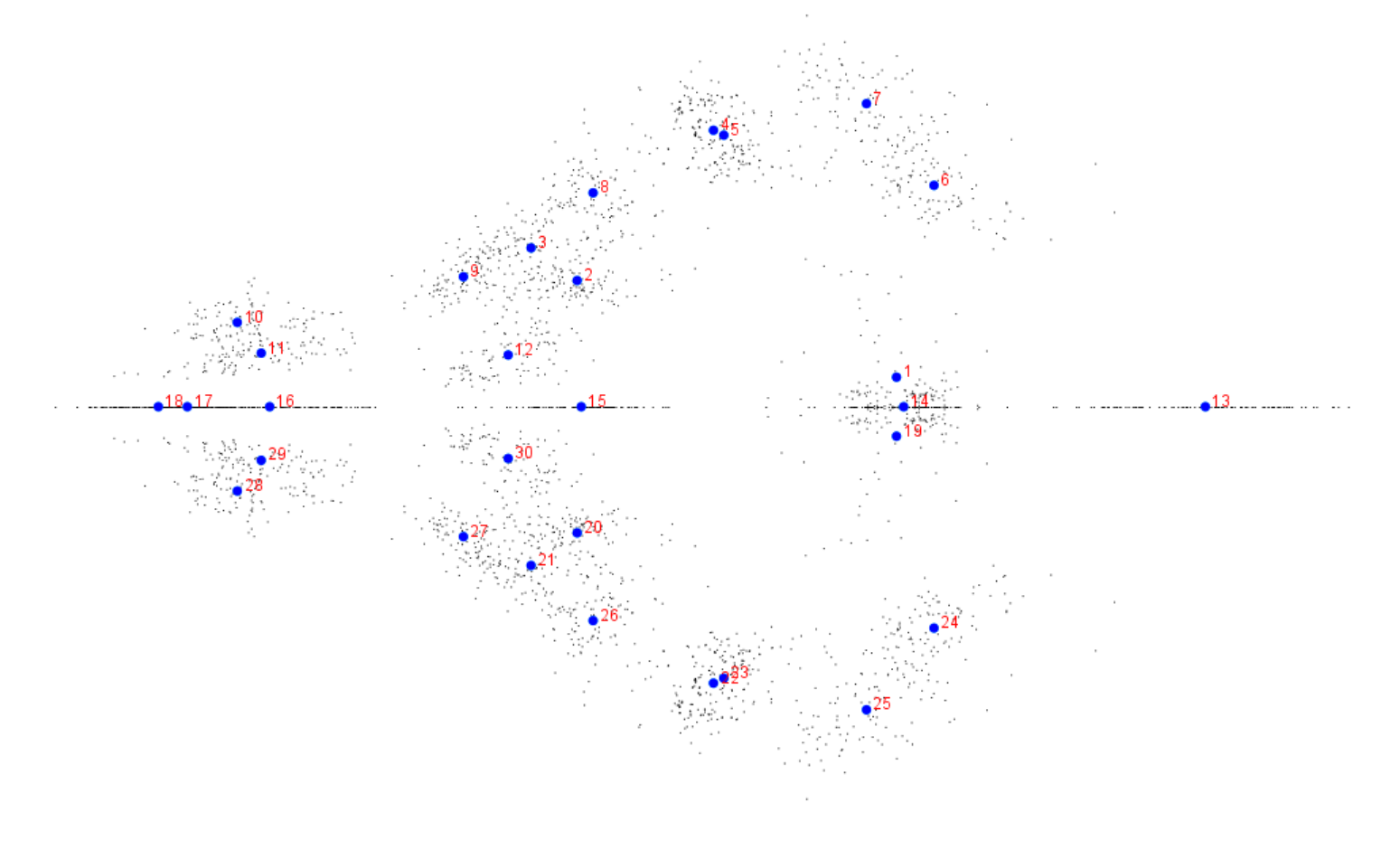
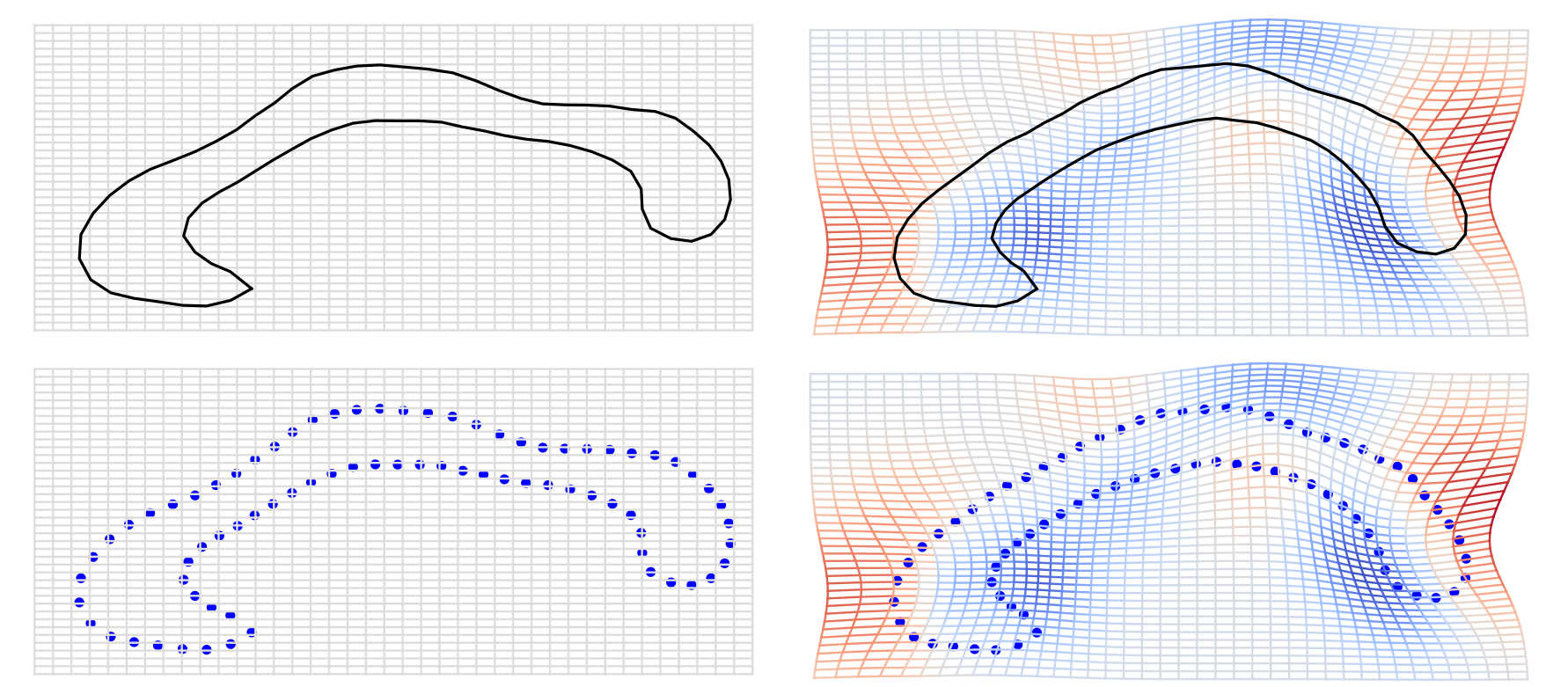
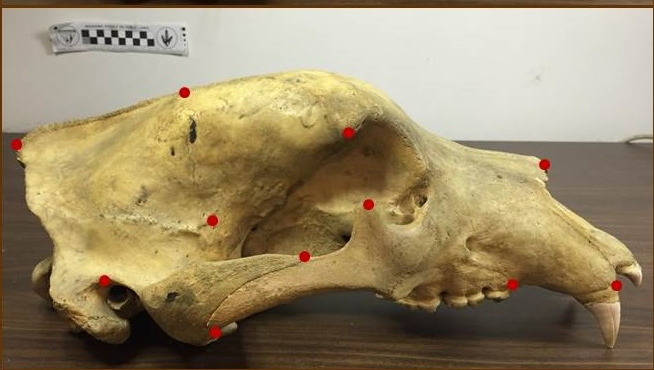
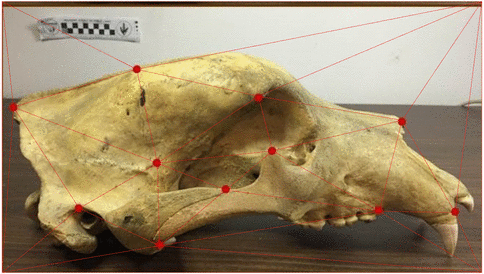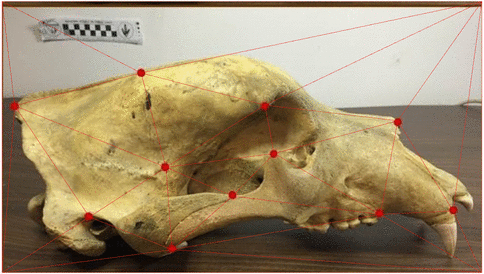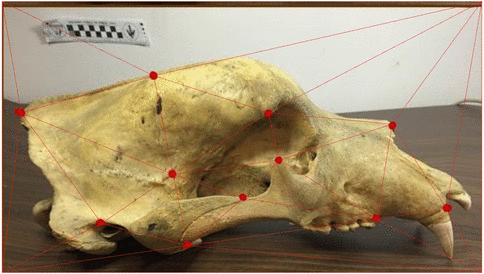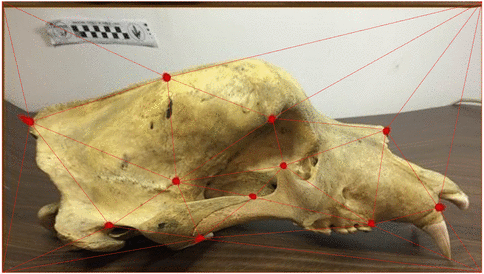
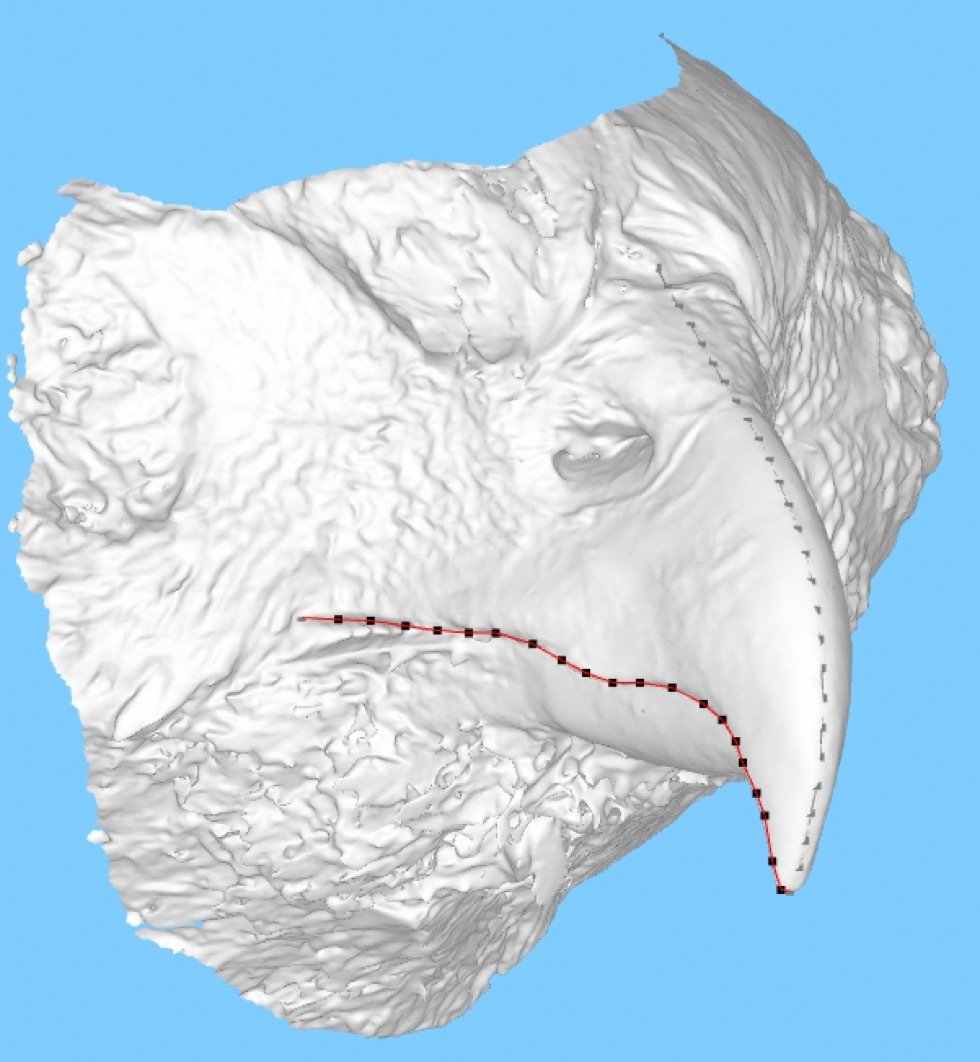
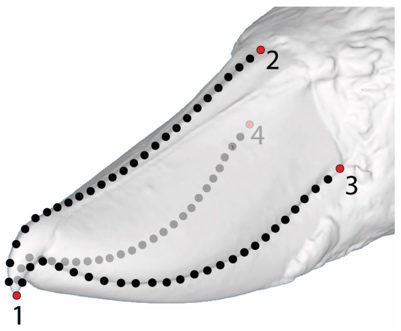
Use of landmarks.
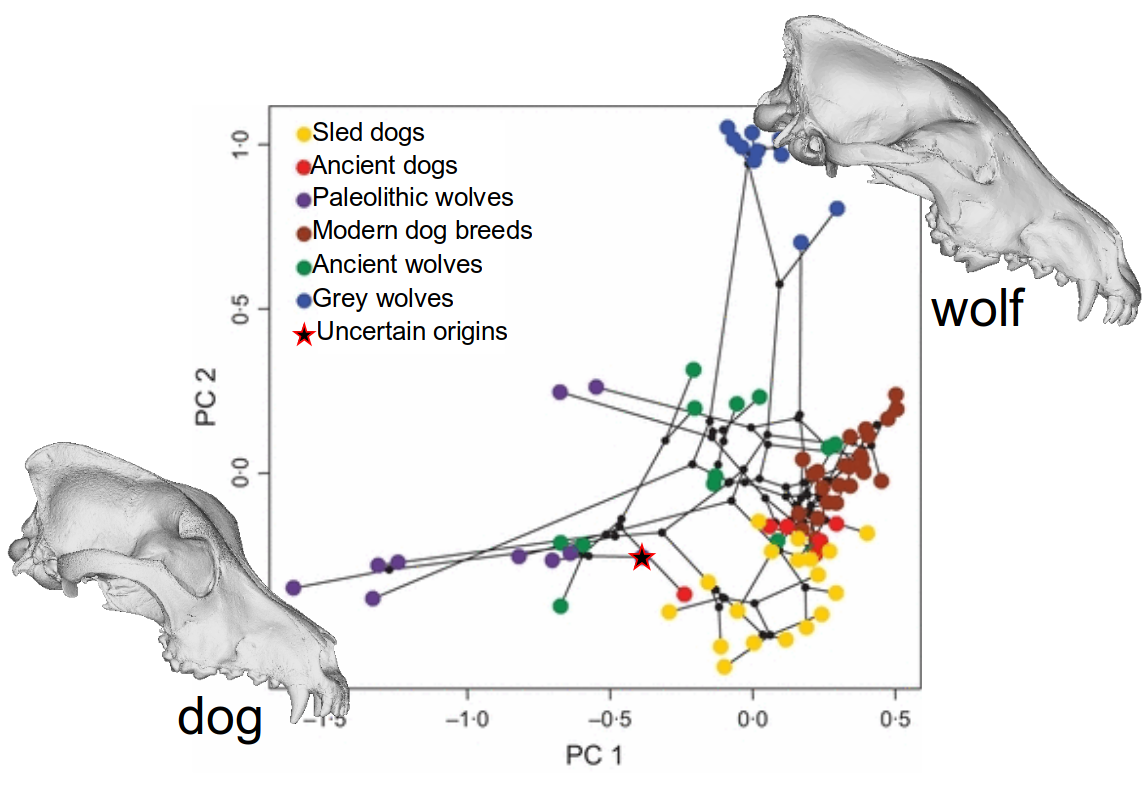
Procrustes alignment.
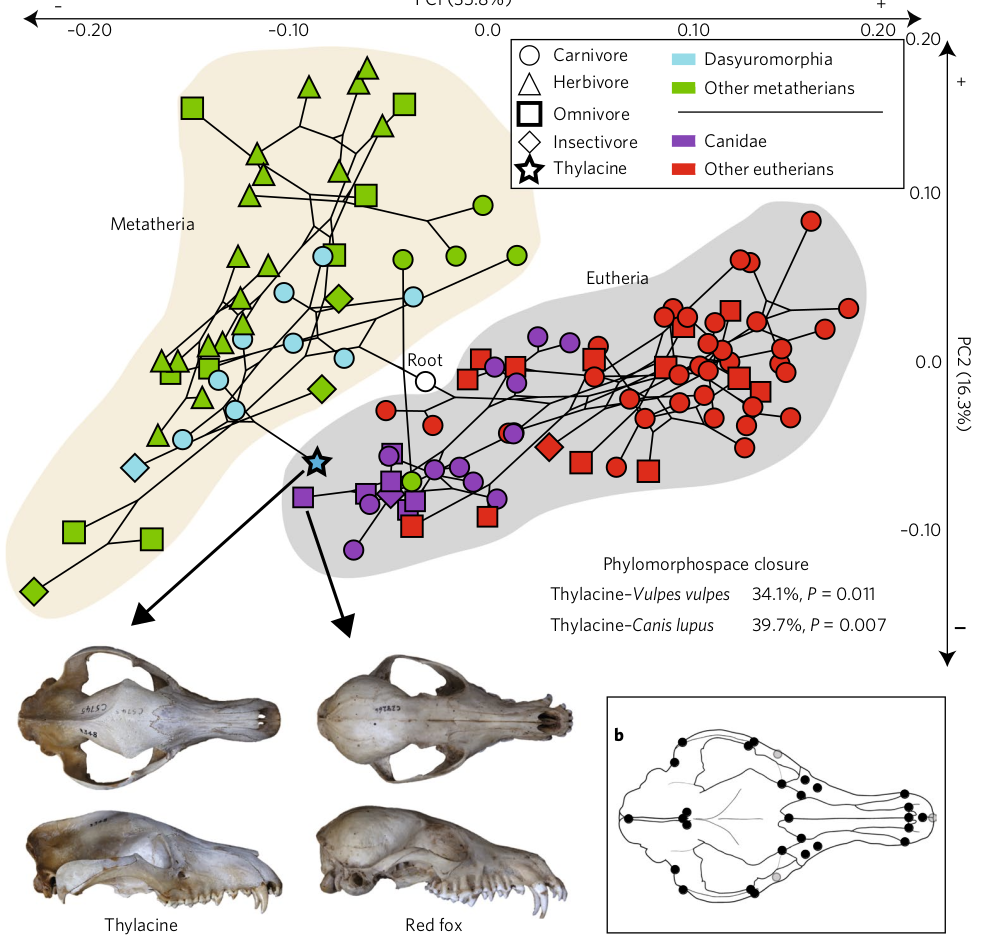
PCA analyses.
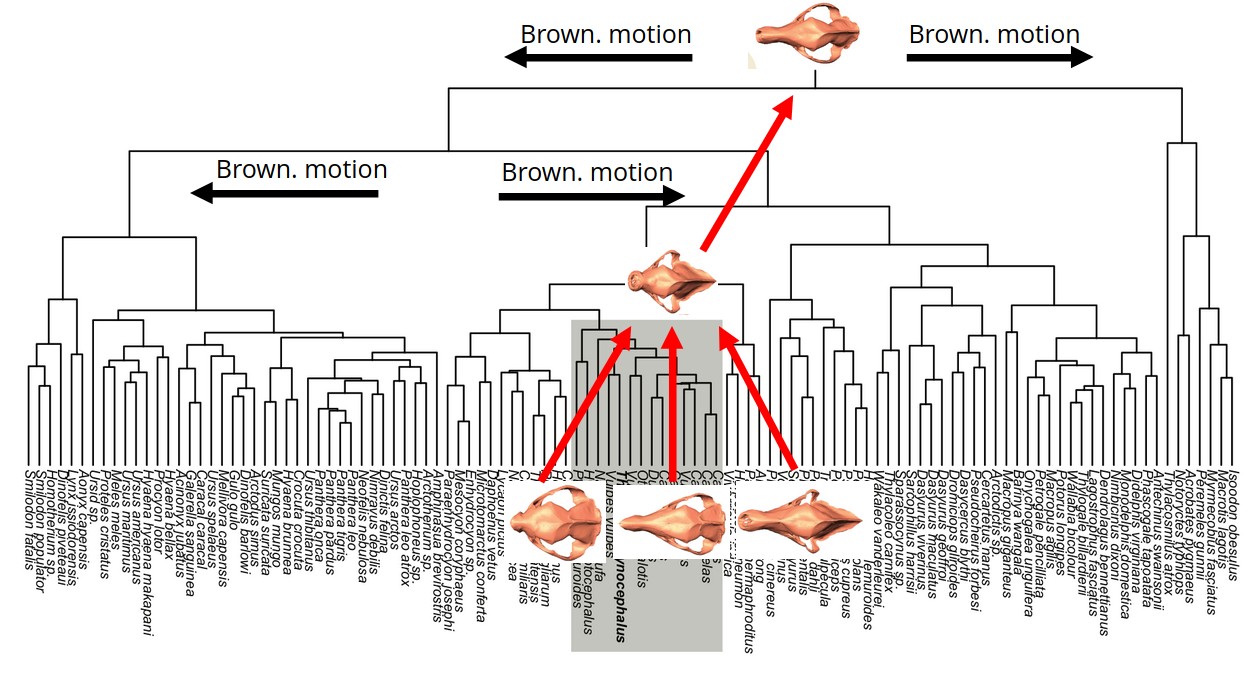
Assume top PCs evolve according to a Brownian motion process.
Then use of methods similar to DNA analyses for computation.
Challenges:
Loss of information in the use of landmarks
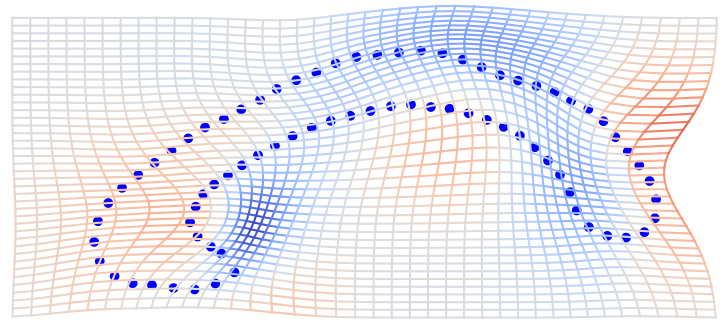
Loss of information in linearization of nonlinear shape space
Loss of information in only using the top PCs
Lack of biological interpretability and justification.
Lack of modeling flexibility.
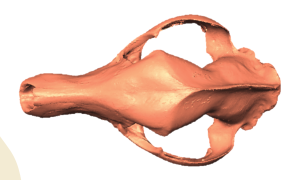
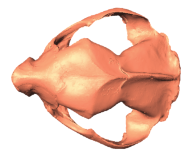
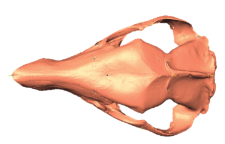
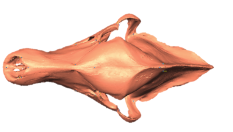
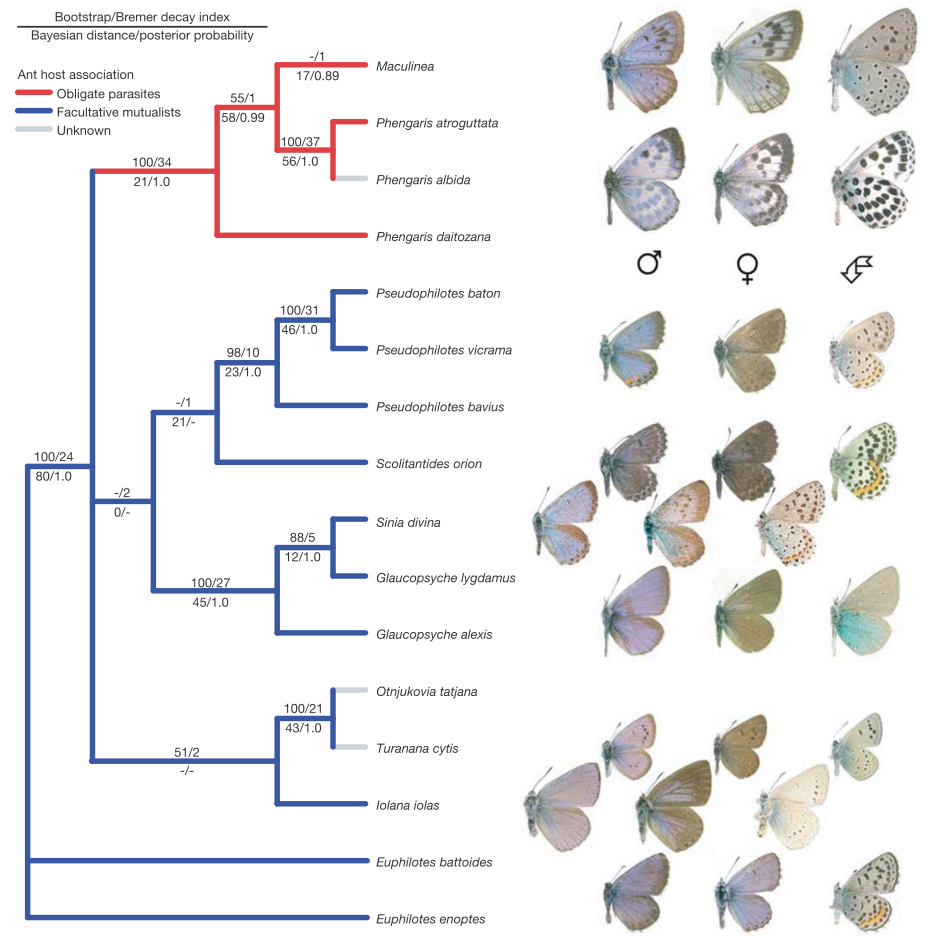
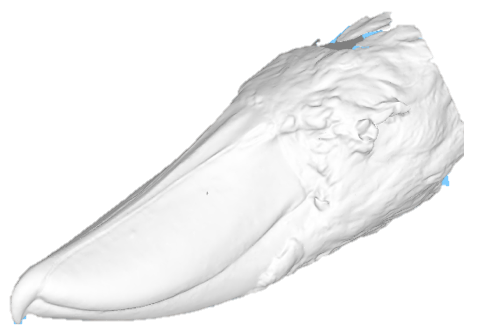
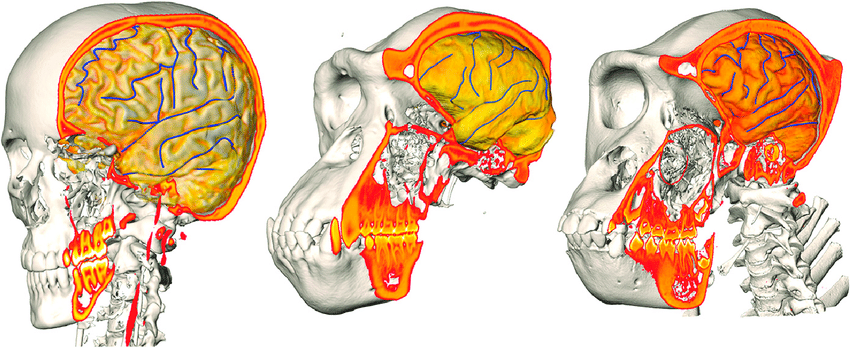
State-of-the-art: full shape is reduced to selected landmarks
State-of-the-art: We have high-res digital morphology images but we cannot use the full shape information
\( \phi \) warp of domain \(\Omega\) (2D or 3D space)
landmarks: \(s=(x_1,\ldots,x_n)\) curves: \(s: \mathbb S^1\to\mathbb R^2\)
surfaces: \(s: \mathbb S^2\to\mathbb R^3\)
\( \phi \)
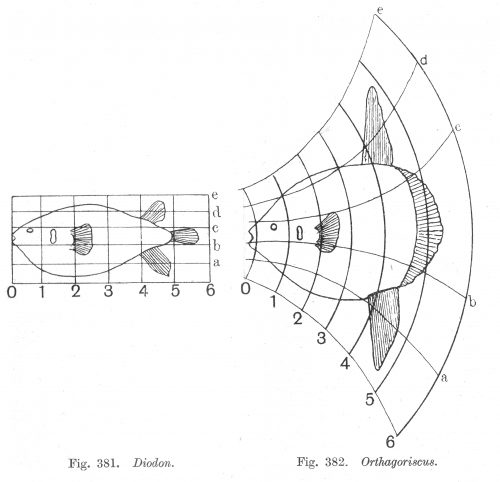
On growth and form, 1917
D'Arcy Thompson
shape \(s_0\)
shape \(s_1\)
stoch. evolution \(s_0\rightarrow s_1\)
Riemannian Brownian motion:
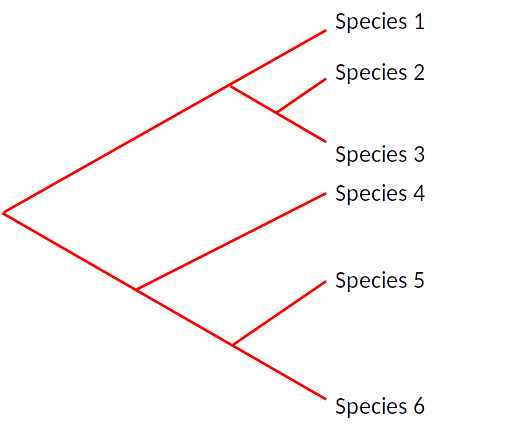
inferring the laws of morphological change
Brown. motion
Brown. motion
Brown. motion
Brown. motion
branch (independent children)
incorporate leaf observations \(x_{V_T}\) into probabilistic model:
\(p(X_t|x_{V_T})\)
Doob’s h-transform
\(h_s(x)=\prod_{t\in\mathrm{ch(s)}}h_{s\to t}(x)\)
conditioned process \(X^*_t\)
approximations \(\tilde{h}\)
guided process \(X^\circ_t\)
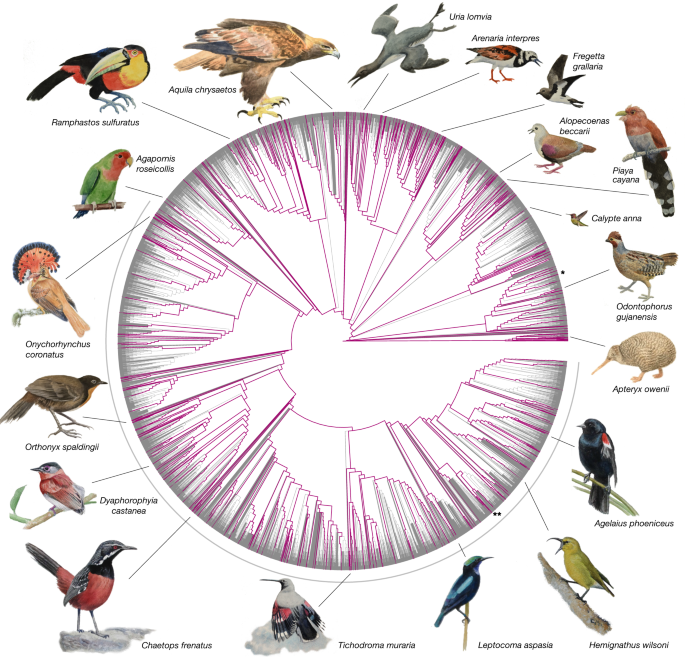
4 cases: Phylogenetics and morphology is foundation for much of bioscience
Rasmus Nielsen
GLOBE UCPH, Berkeley
Phylogenetics
Stefan Sommer
DIKU UCPH
Shape analysis
Christy Hipsley
BIO UCPH
Morphology
Mads Nielsen
DIKU UCPH
Image analysis
Anders Jordan
Natural History Museum of Denmark
Tom Gilbert
GLOBE
Frido Welker
GLOBE
Guojie Zhang
UCPH BIO
Elizabeth Baker, Sofia Stroustrup, Marcus Teller, Lili Bao, Gefan Yang, Liwei Hu, Michael Severinsen, Chao Zhang,
Christine Sarah Andersen + more to come
Josefin Stiller
UCPH BIO
By Stefan Sommer




